Tạo dòng và bước đầu đánh giá mức độ biểu hiện của Hcp1 và AhpC tái tổ hợp của Burkholderia pseudomallei trong Escherichia coli
Trung tâm Nhiệt đới Việt - Nga, Hà Nội, Việt Nam
Viện Y sinh Nhiệt đới, Trung tâm Nhiệt đới Việt – Nga, Số 63 Nguyễn Văn Huyên, Nghĩa Đô, Hà Nội
Email: hoang.dang.hieu.ls@gmail.com
Nội dung chính của bài viết
Tóm tắt
Bệnh melioidosis là bệnh nhiễm trùng nguy hiểm do vi khuẩn Burkholderia pseudomallei gây ra. Các biểu hiện lâm sàng của bệnh không đặc hiệu, khiến việc chẩn đoán sớm medioidosis gặp nhiều khó khăn và dễ dẫn đến điều trị chậm trễ, làm tăng nguy cơ tử vong. Các xét nghiệm huyết thanh học sử dụng kháng nguyên tái tổ hợp được xem là hướng tiếp cận tiềm năng nhờ khả năng phát hiện nhanh kháng thể đặc hiệu kháng B. pseudomallei. Nhiều protein của vi khuẩn này đã được khảo sát nhằm phát triển kháng nguyên phục vụ chẩn đoán melioidosis. Trong số đó, Hcp1 và AhpC được đặc biệt quan tâm nhờ khả năng phản ứng đặc hiệu với kháng thể trong huyết thanh bệnh nhân. Nghiên cứu này nhằm tạo dòng và biểu hiện các gen Hcp1, AhpC, có kích thước lần lượt 510 bp và 549 bp, trong hệ biểu hiện Escherichia coli sử dụng vector pET32a(+). Vector mang cấu trúc dung hợp TrxA-His tag, nhằm cải thiện độ hòa tan của protein và tạo thuận lợi cho quá trình tinh sạch. Kết quả phân tích bằng SDS-PAGE và Western blot xác nhận sự biểu hiện của hai protein đích với kích thước tương ứng khoảng 36 kDa và 38 kDa. Khảo sát tính hòa tan ghi nhận sự khác biệt rõ rệt giữa hai protein: AhpC hiện diện chủ yếu ở dạng hòa tan, trong khi Hcp1 tập trung phần lớn ở phân đoạn không tan. Các kết quả này khẳng định tính khả thi của hệ thống biểu hiện, đồng thời nhấn mạnh sự cần thiết phải tối ưu điều kiện nuôi cấy để nâng cao khả năng thu nhận protein Hcp1 dạng hòa tan, tạo nền tảng cho các bước tinh sạch và phát triển kháng nguyên tái tổ hợp phục vụ cho chẩn đoán trong các nghiên cứu tiếp theo.
Abstract
Cloning and preliminary evaluation of recombinant Hcp1 and AhpC expression from Burkholderia pseudomallei in Escherichia coli
Melioidosis is a severe infectious disease caused by Burkholderia pseudomallei. Nonspecific clinical manifestations make early diagnosis of melioidosis challenging and may lead to delayed treatment, thereby increasing the risk of mortality. Serological assays using recombinant antigens are considered a promising approach due to their ability to rapidly detect antibodies specific to B. pseudomallei. Several proteins of this bacterium have been investigated for the development of diagnostic antigens for melioidosis. Among them, Hcp1 and AhpC have attracted particular attention due to their strong and specific reactivity with antibodies in the serum of patients with melioidosis. This study aimed to clone and express the Hcp1 and AhpC genes, with sizes of 510 bp and 549 bp, respectively, in an Escherichia coli expression system using the pET32a(+) vector. The vector contains a TrxA–His tag fusion construct to improve protein solubility and facilitate purification. SDS-PAGE and Western blot analyses confirmed the expression of the target proteins, with estimated molecular weights of approximately 36 kDa and 38 kDa, respectively. Solubility assessment revealed a marked difference between the two recombinant proteins: AhpC was predominantly present in the soluble fraction, whereas Hcp1 was largely confined to the insoluble fraction. These findings confirm the feasibility of the expression system and highlight the need to optimize culture conditions to improve the recovery of soluble Hcp1, providing a basis for subsequent purification and the development of recombinant antigens for diagnostic applications.
Keywords: AhpC; Burkholderia pseudomallei; Hcp1; melioidosis; recombinant.
Từ khóa
AhpC, Burkholderia pseudomallei, Hcp1, melioidosis, tái tổ hợp
Chi tiết bài viết

Bài báo này được cấp phép theo Creative Commons Attribution 4.0 International License.
Điểm nổi bật:
Hai kháng nguyên Hcp1 và AhpC của Burkholderia pseudomallei được biểu hiện thành công trong E. coli dưới dạng protein tái tổ hợp, mức độ biểu hiện và tính đặc hiệu của protein được xác nhận bằng phân tích SDS-PAGE kết hợp Western blotting.
Đánh giá bước đầu về tính tan cho thấy AhpC chủ yếu hiện diện ở phân đoạn hòa tan, trong khi Hcp1 thu hồi được chiếm ưu thế tuyệt đối ở phân đoạn không tan.
1. ĐẶT VẤN ĐỀ
Melioidosis còn gọi là bệnh Whitmore là bệnh nhiễm trùng nguy hiểm do B. pseudomallei gây ra, tỷ lệ tử vong của bệnh có thể lên tới hơn 40% nếu việc điều trị diễn ra muộn [1]. Bắc Úc và khu vực Đông Nam Á là nơi ghi nhận phần lớn các trường hợp melioidosis trên toàn cầu [2]. Tại Việt Nam, các trường hợp melioidosis thường gia tăng mạnh sau các đợt mưa lớn và lũ lụt. Điển hình, trong đợt mưa lũ cuối năm 2020 tại khu vực miền Trung, số ca mắc đã tăng từ một vài trường hợp lên đến hàng chục trường hợp mỗi tháng [3]. Trong những năm gần đây, do chưa có hệ thống giám sát thống nhất trên phạm vi toàn quốc, số liệu về bệnh chưa được ghi nhận đầy đủ. Tuy nhiên, các nghiên cứu hồi cứu tại một số cơ sở y tế cho thấy bệnh nhân melioidosis vẫn được phát hiện tại nhiều tỉnh, thành phố, với số ca ghi nhận có xu hướng tăng theo thời gian [4,5]. Melioidosis là bệnh khó chẩn đoán do các biểu hiện của bệnh không đặc hiệu, thường tương đồng với lao, sốt rét hoặc các dạng nhiễm khuẩn huyết khác [1]. Nuôi cấy vi sinh được xem là tiêu chuẩn vàng cho phát hiện melioidosis nhưng quy trình xét nghiệm mất nhiều thời gian và kết quả không chính xác với những trường hợp đã sử dụng kháng sinh, trong khi các kỹ thuật PCR lại đòi hỏi trang thiết bị chuyên sâu, hiện đại. Do đó, các xét nghiệm huyết thanh học phát hiện kháng thể đặc hiệu được xem là hướng tiếp cận phù hợp cho sàng lọc và hỗ trợ chẩn đoán [6]. Với thời gian cho kết quả ngắn (từ vài chục phút đến vài giờ), chi phí hợp lý và quy trình kỹ thuật tương đối đơn giản, phương pháp này có thể được triển khai hiệu quả trong các khảo sát diện rộng với quy mô lớn [7, 8]. Việc lựa chọn kháng nguyên phù hợp là yếu tố then chốt, quyết định khả năng phát hiện chính xác của phương pháp này. Nhiều kháng nguyên được sử dụng trong chẩn đoán B. pseudomallei như capsular polysaccharide (CPS), Hemolysin-coregulated protein 1 (Hcp1), Alkyl hydroperoxide reductase C (AhpC), chaperonin GroEL1, O-polysaccharide (OPS) [9, 10]. Trong đó, Hcp1 và AhpC là hai kháng nguyên có liên quan mật thiết đến cơ chế gây bệnh của B. pseudomallei và có khả năng gây đáp ứng miễn dịch tốt phù hợp để phát triển công cụ dự đoán melioidosis. Hcp1 thuộc hệ thống bài tiết type VI (T6SS) của B. pseudomallei, được ghi nhận có liên quan đến quá trình nhiễm trùng của B. pseudomallei và gia tăng biểu hiện trong điều kiện này [11]. AhpC là enzyme chống oxy hóa tham gia bảo vệ vi khuẩn trước stress oxy hóa của vật chủ [12]. Hcp1 và AhpC đã được nhiều nghiên cứu sử dụng làm kháng nguyên trong phát hiện B. pseudomallei. Nghiên cứu của Wagner và cộng sự tại Thái Lan cho thấy xét nghiệm huyết thanh đa kháng nguyên giúp tăng độ nhạy trong chẩn đoán. Tại Việt Nam, nhóm nghiên cứu của Trần Thị Lệ Quyên cũng chứng minh hiệu quả của Hcp1, AhpC trong việc phát hiện B. pseudomallei bằng ELISA [8, 10]. Việc sử dụng kháng nguyên Hcp1, AhpC trong việc phát hiện melioidosis cho thấy độ nhạy và độ đặc hiệu còn biến động lớn giữa các quần thể nghiên cứu, đặc biệt bị ảnh hưởng bởi nền miễn dịch ở vùng lưu hành. Ngoài ra, việc thiếu chuẩn hóa quy trình và ngưỡng đánh giá khiến kết quả chưa đồng nhất [13]. Trong bối cảnh đó, việc chủ động tạo nguồn kháng nguyên chất lượng trong nước có ý nghĩa quan trọng, giúp giảm phụ thuộc vào nguồn nhập khẩu, đảm bảo tính ổn định và phù hợp với đặc điểm dịch tễ địa phương. Đồng thời, đây cũng là cơ sở để chuẩn hóa quy trình xét nghiệm và phát triển các phương pháp chẩn đoán hiệu quả, phù hợp với điều kiện thực tiễn
Nghiên cứu này trình bày kết quả tạo dòng và bước đầu đánh giá mức độ biểu hiện của protein Hcp1 và AhpC tái tổ hợp trong tế bào E. coli BL21, nhằm tạo nguồn kháng nguyên phục vụ phát triển các phương pháp chẩn đoán B. pseudomallei.
2. VẬT LIỆU VÀ PHƯƠNG PHÁP
2.1 Vật liệu
2.1.1. Trình tự mồi sử dụng
Các cặp mồi trong nghiên cứu được liệt kê tại Bảng 1. Trình tự được gạch chân là vị trí nhận diện của các enzyme NcoI và XhoI. Trình tự gen Hcp1 (NZ_AP028078.1) và AhpC (CP002833.1) của vi khuẩn B. pseudomallei trên ngân hàng Genbank được tham khảo cho quá trình thiết kế mồi.
Bảng 1. Mồi nhân bản gen Hcp1, AhpC
STT | Tên mồi | Trình tự mồi 5’-3’ | Gen đích | Kích thước (bp) |
1 | Fw_Hcp1 (Nco I) | 5’-GCCATGGATATGCTGGCCGGAATATATCTC-3’ | Hcp1 | 510 |
2 | Rw_Hcp1 (Xho I) | 5’-GCTCGAGTCAGCCATTCGTCCAGTTTG-3’ | ||
3 | Fw_AhpC (Nco I) | 5’-GCCATGGATATGAAGACCGTGGGCGATAAAC-3’ | AhpC | 549 |
4 | Rv_AhpC (Xho I) | 5’-GCTCGAGTTACAGCGTCGCGCCGCCGAT-3’ |
2.1.2. Các chủng vi khuẩn và vector biểu hiện
Các chủng vi khuẩn và vector sử dụng được cung cấp bởi Phòng Độc học và Các bệnh nhiệt đới, Trung tâm Nhiệt đới Việt - Nga. Cụ thể:
- Chủng vi khuẩn chủ: Escherichia coli DH5α (Thermo Scientifi - Mỹ), chủng Escherichia coli BL21 (DE3) (Thermo Scientifi - Mỹ).
- DNA khuôn: DNA tổng số từ chủng vi khuẩn B. pseudomallei, được lấy từ nguồn mẫu của đề tài hợp tác Việt - Nga giai đoạn 2016–2020.
- Vector biểu hiện: pET-32a(+) (Novagen - Mỹ).
2.1.3. Hóa chất và trang thiết bị
Hóa chất và môi trường: Chất cảm ứng Isopropyl β-D-1-thiogalactopyranoside (IPTG) (Meridian Bioscience, Mỹ), môi trường LB (chứa 10 g/L peptone (Himedia - Ấn độ), 5 g/L cao nấm men (Himedia - Ấn độ), 10 g/L NaCl (Biobasic - Canada), pH 7,0, kháng sinh ampicillin (Biobasic - Canada). Kit tinh sạch sản phẩm PCR, Gel (ABT - Việt Nam), enzym cắt giới hạn NcoI, XhoI (Thermo - Lithuania), Enzyme T4 DNA ligase (Vazyme - Trung Quốc), Kit tách chiết và tinh sạch plasmid (Genedirex - Đài Loan), Master mix 2X (Vazyme - Trung Quốc).
Các hóa chất phục vụ điện di DNA, protein bao gồm: Thang chuẩn DNA (Genedirex - Đài Loan), Đệm TAE (ABT - Việt Nam), Marker DNA (Genedirex - Đài Loan), Tris-base (ABT, Việt Nam), Acrylamide (Biobasic - Canada), bis-acrylamide (Serva, Đức), thuốc nhuộm Coomassie blue (Biobasic - Canada), methanol (Merck - Đức), acid acetic (Biobasic - Canada), SDS (Biobasic - Canada), TEMED (Biobasic - Canada), APS (Merck - Đức), glycine (Sigma, Mỹ), kháng thể anti - Histag(R&D Systems - Mỹ), kháng thể kháng IgG chuột liên hợp HRP (R&D Systems - Mỹ) và thang chuẩn protein (Thermo, Mỹ), TBST 1X (20 mM Tris-HCl pH 7,5, 150 mM NaCl, 0,1% Tween-20).
Thiết bị: Thí nghiệm lên men được tiến hành trên tủ lắc gia nhiệt NB-205VL (N-Biotek, Hàn Quốc). Các mẫu DNA và protein sau đó được quan sát và phân tích bằng hệ thống điện di ngang, điện di đứng và thiết bị chụp ảnh Gel Doc EZ Imager (Bio-Rad, Mỹ).
2.2. Phương pháp nghiên cứu
2.2.1. Tạo vector pET32a(+) mang gen Hcp1/AhpC
Các gen Hcp1 và AhpC được thu nhận và khuếch đại từ DNA tổng số B. pseudomallei bằng phản ứng PCR. Phản ứng PCR được thực hiện trong tổng thể tích 25 µL, bao gồm 12,5 µL dung dịch 2X PCR Master Mix, 1 µL (mồi xuôi và mồi ngược nồng độ cuối 0,2 µM), 2 µL DNA khuôn và bổ sung nước không chứa nuclease cho đủ thể tích. Chu trình nhiệt được thiết lập với bước biến tính ban đầu ở 95°C trong 3 phút, tiếp theo là 30 chu kỳ gồm: biến tính ở 95°C trong 30 giây, gắn mồi ở 56°C trong 30 giây và kéo dài ở 72°C trong 40 giây. Sau đó, thực hiện bước kéo dài cuối ở 72°C trong 5 phút, và giữ ở 4 – 12°C cho đến khi thu mẫu. Các cặp mồi sử dụng trong nghiên cứu được trình bày tại Bảng 1.
Vector pET-32a(+) và đoạn DNA khuếch đại được cắt bằng enzyme giới hạn NcoI và XhoI. Phản ứng cắt enzyme giới hạn được thực hiện trong tổng thể tích 50 µL, bao gồm 5 µL dung dịch đệm 10X, khoảng 0,5 – 1 µg DNA plasmid, 1 µL mỗi enzyme giới hạn (NcoI và XhoI) và bổ sung nước không chứa nuclease cho đủ thể tích. Hỗn hợp được ủ ở 37°C trong 2 giờ nhằm đảm bảo quá trình cắt diễn ra hoàn toàn.
Sản phẩm cắt enzyme sau đó được tinh sạch và nối với nhau bằng T4 DNA ligase. Tiếp theo được chuyển vào E. coli DH5α bằng sốc nhiệt, sau đó tiến hành sàng lọc ở 37°C trên môi trường thạch LB có mặt của kháng sinh ampicillin (100 µg/mL) (LBA) qua đêm. Các khuẩn lạc thu được được nuôi tăng sinh và tách chiết plasmid bằng bộ kit miniPREP (Genedirex - Đài Loan). Plasmid sau đó được kiểm tra bằng PCR với mồi đặc hiệu tương ứng. Những mẫu dương tính được giải trình tự và ký hiệu lần lượt là pET32a-Hcp1 và pET32a-AhpC.
2.2.2. Biểu hiện Hcp1/AhpC trong tế bào E. coli BL21
Các plasmid pET32a-Hcp1 và pET32a-AhpC được biến nạp vào Escherichia coli BL21 bằng phương pháp sốc nhiệt, sau đó các dòng tế bào mang plasmid tái tổ hợp được tuyển chọn trên môi trường LBA. Chủng E. coli BL21 mang gen mã hóa RNA polymerase T7 trong hệ gen, cho phép cảm ứng biểu hiện protein đích thông qua promoter T7 trên vector pET khi bổ sung IPTG [14]. Các khuẩn lạc đơn đại diện được lựa chọn, nuôi tăng sinh và bảo quản ở −80°C trong dung dịch glycerol 30% nhằm duy trì khả năng sống và tính ổn định của các chủng mang plasmid trong quá trình lưu giữ dài hạn, đồng thời cung cấp nguồn tế bào phục vụ cho các thí nghiệm tiếp theo [15].
2.2.3. Khảo sát biểu hiện của Hcp1 và AhpC trong E. coli BL21
Tiến hành nuôi các dòng tế bào biểu hiện Hcp1, AhpC ở 37°C trong 5–10 mL môi trường LBA qua đêm, lắc 220 vòng/phút. Sau đó chuyển dịch nuôi sang môi trường LBA mới theo tỷ lệ 1:20 (v/v), tiếp tục nuôi lắc cho đến khi OD₆₀₀ đạt 0,6–0,8. Bổ sung IPTG 0,5 Mm để cảm ứng tăng khả năng biểu hiện và tiếp tục nuôi lắc trong 16 giờ ở 37oC. Tiến hành ly tâm lạnh (5000 rpm, 10 phút) để thu sinh khối, sau đó hòa tan trong đệm PBS 1X. Quá trình phá màng tế bào được thực hiện bằng siêu âm trên đá 40 lần ở tần số 30 Hz, xung 15 giây, nghỉ 10 giây để thu nhận protein. Sau đó, mẫu được ly tâm ở 12.000 vòng/phút trong 10 phút ở 4°C để tách pha tan và không tan; phần dịch nổi được thu để đánh giá protein pha tan, trong khi phần cặn được hòa tan lại trong đệm PBS 1X để thu protein pha không tan. Dịch protein được phân tích bằng điện di SDS-PAGE trên gel polyacrylamide 12,6% và nhuộm Coomassie Brilliant Blue nhằm đánh giá mức độ biểu hiện protein.
Kỹ thuật Western blotting được áp dụng nhằm xác định khả năng nhận diện đặc hiệu giữa protein và kháng thể. Sau điện di, protein được chuyển lên màng PVDF bằng thiết bị chuyển màng của Bio-Rad (Mỹ) ở 100 V trong 90 phút. Màng sau đó được rửa bằng TBST 1X, và tiến hành blocking bằng dung dịch sữa tách béo 5% pha trong TBST 1X với thời gian 1 giờ. Tiếp theo ủ màng qua đêm ở 4°C với kháng thể sơ cấp kháng His-tag nguồn gốc thỏ (pha loãng 1:10.000 trong TBST 1X), sau đó rửa bằng TBST 1X và tiếp tục ủ trong 1 giờ với kháng thể thứ cấp kháng IgG thỏ liên hợp HRP (pha loãng 1:10.000 trong TBST 1X) ở nhiệt độ phòng. Tiến hành phát hiện protein bằng việc bổ sung cơ chất TMB, cho phép ghi nhận sự xuất hiện của các băng protein Hcp1 và AhpC trên màng lai.
3. KẾT QUẢ
3.1. Tạo cấu trúc vector pET32a(+)-AhpC/Hcp1
Kết quả điện di trên gel agarose cho thấy một băng DNA duy nhất, rõ nét và trùng với kích thước lý thuyết của gen Hcp1, AhpC. Không có băng tín hiệu DNA nào ở mẫu đối chứng âm (không chứa DNA khuôn) được phát hiện. Những kết quả này khẳng định gen Hcp1 và AhpC đã được nhân bản thành công từ DNA của vi khuẩn B. pseudomallei (Hình 1).
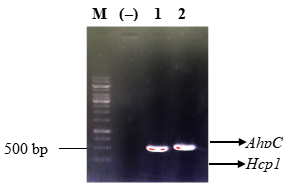
Hình 1. Kết quả PCR gen Hcp1, AhpC từ DNA vi khuẩn B. pseudomallei
M: Marker DNA 1kb; (-) đối chứng âm không chứa DNA vi khuẩn B. pseudomallei; 1: gen Hcp1; 2: gen AhpC
Các gen đích sau khuếch đại được gắn với vector pET32a(+) tại hai vị trí của enzyme NcoI và XhoI, hình thành plasmid tái tổ hợp pET32a-Hcp1 và pET32a-AhpC. Hai cấu trúc plasmid này được biến nạp vào E. coli DH5α nhằm phục vụ quá trình nhân bản và lưu giữ vector chứa gen đích.
Sau quá trình biến nạp, các khuẩn lạc riêng rẽ được lựa chọn và sử dụng để nuôi cấy, từ đó thu nhận plasmid phục vụ các phân tích tiếp theo. Gen Hcp1 và AhpC trong plasmid được xác định thông qua PCR với các cặp mồi đặc hiệu cho Hcp1 và AhpC. Hình ảnh điện di trên gel agarose ghi nhận hai băng DNA có kích thước hơn 500 bp, trùng với các mẫu chứng dương (DNA tổng số của B. pseudomallei). Trong khi đó, plasmid pET32a(+) không mang gen chèn sử dụng làm đối chứng âm không xuất hiện sản phẩm khuếch đại (Hình 2). Những kết quả này xác nhận việc gắn thành công các gen Hcp1 và AhpC vào vector pET32a(+).
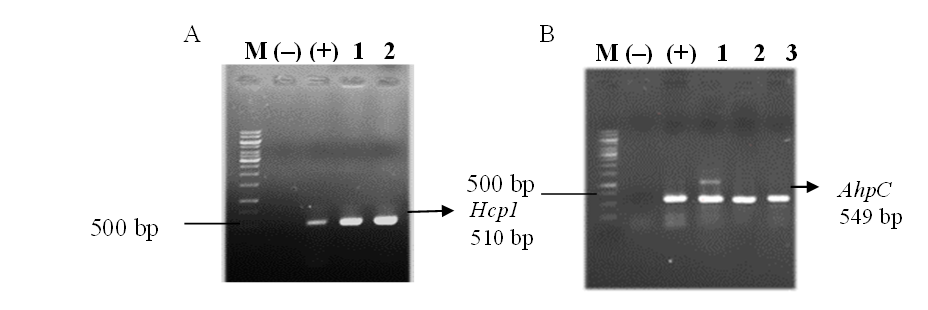
Hình 2. Kết quả điện kiểm tra plasmid mang gen Hcp1 (A) và AhpC (B) M: Marker 1kb; (-): chứng âm; (+): chứng dương; 1-3: plasmid tách từ các khuẩn lạc chọn lọc
Các plasmid cho tín hiệu PCR dương tính được xác nhận bằng giải trình tự Sanger. Kết quả đối chiếu cho thấy trình tự gen Hcp1 thu nhận từ plasmid tái tổ hợp trùng khớp hoàn toàn với trình tự tương ứng của B. pseudomallei trên GenBank (NZ_AP028078.1) (Hình 3A). Đối với gen AhpC, phân tích trình tự ghi nhận sự thay thế nucleotide A bằng G tại vị trí 84 so với trình tự tham chiếu CP002833.1. Mặc dù vậy, sự thay thế nucleotide này không ảnh hưởng đến trình tự axit amin dịch mã của protein AhpC (Hình 3B).

Hình 3. Kết quả kiểm tra trình tự gen Hcp1 (A); AhpC (B) trong vector tái tổ hợp bằng giải trình tự Sanger
3.2. Tạo chủng biểu hiện mang plasmid pET32a-Hcp1/AhpC trong E. coli BL21
Chủng E. coli BL21 được sử dụng làm hệ thống biểu hiện sau khi tiếp nhận các plasmid tái tổ hợp pET32a-Hcp1 và pET32a-AhpC. Sau biến nạp, xuất hiện các khuẩn lạc tách biệt được quan sát trên môi trường chọn lọc LBA, cho thấy tế bào E. coli BL21 đã tiếp nhận plasmid tái tổ hợp thành công (Hình 4). Một số khuẩn lạc đơn được lựa chọn để tăng sinh và lưu trữ ở −80°C trong môi trường LBA lỏng bổ sung 30% glycerol.
Hình 4. Tạo dòng E. coli BL21 chứa plasmid tái tổ hợp. (A): Kết quả biến nạp vector pET 32a-Hcp1; (B): Kết quả biến nạp vector pET 32a-AhpC
3.3. Đánh giá mức độ biểu hiện của AhpC và Hcp1 trong E. coli BL21 mang vector pET32a-Hcp1 và pET32a-AhpC
Phân tích SDS-PAGE cho thấy sự xuất hiện của các băng protein có cường độ cao tại vùng khối lượng phân tử lớn hơn 35 kDa ở các mẫu biểu hiện protein tái tổ hợp, trong khi các băng tương ứng không được phát hiện ở mẫu đối chứng mang vector pET32a(+) không chứa gen chèn (Hình 5A). Kết quả này cho thấy sự biểu hiện thành công của các protein đích trong hệ thống biểu hiện đã thiết kế.
Để xác nhận thêm tính đặc hiệu của các protein thu được, các mẫu được phân tích bằng Western blotting sử dụng kháng thể kháng His-tag. Kết quả cho thấy các tín hiệu đặc hiệu xuất hiện tại vị trí khoảng 36 kDa và 38 kDa, tương ứng với các protein dung hợp Hcp1 và AhpC. Ngược lại, mẫu đối chứng pET32a(+) chỉ ghi nhận tín hiệu tại khoảng 18 kDa và không xuất hiện các băng tín hiệu tại vị trí của protein đích (Hình 5B). Những kết quả này khẳng định sự biểu hiện đặc hiệu của các protein tái tổ hợp mang thẻ His.
Hình 5. Kết quả (A): SDS PAGE; B: western blot
(M: Marker protein; 1: pET 32a(+); 2: pET 32a-Hcp1; 3: pET 32a-AhpC)
Tiếp theo, để đánh giá trạng thái hòa tan của các protein biểu hiện, dịch tế bào sau ly giải được phân tách thành hai phân đoạn hòa tan và không tan và phân tích bằng SDS-PAGE (Hình 6). Kết quả cho thấy protein AhpC chủ yếu hiện diện trong phân đoạn hòa tan, trong khi protein Hcp1 được phát hiện chủ yếu ở phân đoạn kết tủa. Điều này cho thấy AhpC có xu hướng tồn tại ở trạng thái hòa tan tốt hơn so với Hcp1 trong cùng điều kiện biểu hiện đã khảo sát.

Hình 6. Kết quả khảo sát sự phân bố Ahpc và Hcp1 trong các phân đoạn protein (M: Marker protein; TS: Dịch chiết tế bào toàn phần; T: phân đoạn hòa tan; KT: phân đoạn không tan)
4. THẢO LUẬN
Phân tích trình tự của plasmid pET32a-Hcp1 và pET32a-AhpC cho thấy trình tự vùng gen chèn và vùng nối với vector không xuất hiện codon kết thúc sớm và không có hiện tượng lệch khung đọc, trình tự gen Hcp1 thu nhận hoàn toàn trùng khớp với trình tự tham chiếu của B. pseudomallei trên cơ sở dữ liệu GenBank, xác nhận đoạn gen mục tiêu đã được chèn chính xác vào vector biểu hiện, không phát sinh sai lệch nucleotide trong quá trình tạo dòng và nhân bản plasmid. Đối với gen AhpC, phân tích trình tự ghi nhận sự thay thế một nucleotide so với trình tự tham chiếu ban đầu. Tuy nhiên thay đổi này không ảnh hưởng đến bộ ba mã hóa nên không làm thay đổi axit amin cũng như trình tự protein được mã hóa. Ngoài ra kết quả đối chiếu với các trình tự khác trên NCBI cho thấy AhpC có độ tương đồng 100%, chứng tỏ sự thay thế nucleotide quan sát được chỉ là đột biến đồng nghĩa, hiện tượng này được ghi nhận phổ biến trong hệ gen của B. pseudomallei [16]. Kết quả này xác nhận gen Hcp1 và AhpC đã được chèn chính xác vào plasmid pET32a.
Các protein tái tổ hợp Hcp1 và AhpC của B. pseudomallei đã được sử dụng rộng rãi làm kháng nguyên trong các sinh phẩm chẩn đoán melioidosis, đặc biệt trong các xét nghiệm miễn dịch như ELISA và test nhanh. Nhiều nghiên cứu ghi nhận hai protein này có khả năng nhận diện miễn dịch tốt, cho phép phân biệt hiệu quả giữa mẫu huyết thanh bệnh nhân và đối chứng [8,10]. Do đó, việc tạo dòng và biểu hiện thành công các protein này có ý nghĩa quan trọng trong việc cung cấp nguồn nguyên liệu phục vụ chẩn đoán. Trong nghiên cứu này, các protein Hcp1 và AhpC được biểu hiện trong hệ E. coli sử dụng vector pET32a(+). Dưới sự kiểm soát của promoter T7, các protein được biểu hiện dưới dạng dung hợp với thioredoxin (TrxA) và His-tag ở đầu N-terminal, góp phần hỗ trợ phát hiện, tinh sạch và cải thiện độ hòa tan. Các protein dung hợp có khối lượng dự đoán khoảng 36 kDa (Hcp1) và 38 kDa (AhpC). Kết quả SDS-PAGE và Western blot cho thấy các băng protein tương ứng với kích thước dự đoán, xác nhận sự biểu hiện thành công trong E. coli BL21 (Hình 5).
Phân tích tính tan cho thấy hai protein biểu hiện từ cùng hệ pET32a(+) có sự phân bố khác nhau sau ly giải tế bào (Hình 6). AhpC chủ yếu hiện diện ở pha hòa tan, trong khi Hcp1 tập trung ở pha không hòa tan. Mặc dù TrxA được biết đến với vai trò cải thiện độ hòa tan của protein ngoại lai trong E. coli [17,18], và hiệu quả này đã được ghi nhận trong nghiên cứu của Trịnh Văn Toàn [18], kết quả thu được đối với Hcp1 trong nghiên cứu này cho thấy protein vẫn chủ yếu tồn tại ở dạng không tan. Xu hướng này phù hợp với báo cáo của Trần Thị Lệ Quyên (2020), khi Hcp1 được thu nhận chủ yếu dưới dạng thể vùi [19].
Sự khác biệt về phân bố giữa AhpC và Hcp1 gợi ý rằng, trong cùng kiện biểu hiện, khả năng hòa tan của protein có thể chịu ảnh hưởng không chỉ bởi hệ vector mà còn bởi đặc tính cấu trúc nội tại của từng protein. Do đó, cần tiếp tục tối ưu các điều kiện biểu hiện nhằm cải thiện tỷ lệ thu nhận Hcp1 ở dạng hòa tan.
5. KẾT LUẬN
Hai kháng nguyên Hcp1 và AhpC của B. pseudomallei đã được biểu hiện trong E. coli BL21 thông qua vector pET32a(+). Protein tái tổ hợp được xác nhận bằng phân tích SDS-PAGE và Western blot. Phân tích protein sau ly giải cho thấy AhpC ưu thế ở dạng hòa tan, trong khi Hcp1 chủ yếu tồn tại ở phần không hòa tan, cho thấy sự khác biệt về đặc tính nội tại giữa hai protein. Kết quả này cho thấy việc cần thiết phải tối ưu điều kiện biểu hiện để nâng cao hiệu suất thu nhận Hcp1 hòa tan. Những dữ liệu thu được cung cấp cơ sở cho các bước tinh sạch kháng nguyên tái tổ hợp, hướng tới ứng dụng trong chẩn đoán bệnh melioidosis.
Hạn chế của nghiên cứu: Mặc dù các protein tái tổ hợp Hcp1 và AhpC đã được biểu hiện thành công, nghiên cứu này chưa đánh giá hoạt tính kháng nguyên của các protein thu nhận. Ngoài ra, việc biểu hiện dưới dạng dung hợp với thẻ TrxA–His có thể ảnh hưởng đến cấu trúc và che khuất một số epitope, từ đó làm thay đổi khả năng nhận diện miễn dịch so với protein tự nhiên. Do đó, cần có các nghiên cứu tiếp theo nhằm đánh giá tính kháng nguyên và xác nhận tiềm năng ứng dụng của các protein này trong chẩn đoán
Lời cảm ơn: Nghiên cứu này là một phần kết quả từ nhiệm vụ cấp Viện Y sinh Nhiệt đới, Trung tâm Nhiệt đới Việt - Nga “Nghiên cứu tạo protein Hcp1, AhpC đặc hiệu của vi khuẩn Burkholderia pseudomallei bằng công nghệ DNA tái tổ hợp”. Mã số YS.Đ2.07/24.
Tuyên bố về việc sử dụng trí tuệ nhân tạo (AI): Các tác giả khẳng định không sử dụng bất kỳ công cụ trí tuệ nhân tạo nào để tạo ra hoặc chỉnh sửa nội dung khoa học của bản thảo này. Mọi phân tích, diễn giải và kết luận hoàn toàn do các tác giả thực hiện.
Tuyên bố về đóng góp của tác giả: Hoàng Đăng Hiếu: Phương pháp luận, viết bản thảo; Bùi Thị Thanh Nga, Trịnh Văn Toàn, Nguyễn Văn Đoài, Nguyễn Minh Anh, Nguyễn Kim Ngân: Thực hiện thí nghiệm, phân tích và xử lý số liệu; Chử Lương Luân: Phương pháp luận; Võ Viết Cường, Lê Thị Lan Anh: Rà soát và biên tập.
Tuyên bố về xung đột lợi ích: Các tác giả khẳng định không có bất kỳ lợi ích xung đột nào ảnh hưởng đến công trình nghiên cứu được báo cáo trong bài báo này.
Tài liệu tham khảo
- W. J. Wiersinga, et al., Melioidosis, Nature Reviews Disease Primers, Vol. 4, p.17107, 2018 Feb 1. DOI: 10.1038/nrdp.2017.107
- L. Pang, et al., Melioidosis, Singapore, 2003–2014, Emerging Infectious Diseases, Vol. 4, No. 1, pp.140-143, 2018. DOI: 10.3201/eid2401.161449
- Nguyễn Duy Bình, Đặng Thị Hoài Thu và Trần Xuân Chương, Nghiên cứu chùm ca bệnh melioidosis tại miền Trung Việt Nam sau đợt lũ lịch sử năm 2020, Tạp chí Y Dược học, tập 12, số 4, 2022. DOI: 10.34071/jmp.2022.4.16
- Lê Quốc Hũng, Đỗ Thị Ngọc Khánh, Bệnh nhân melioidosis thể nhiễm khuẩn huyết ở một bệnh viện tuyến cuối miền Nam Việt Nam: Đặc điểm lâm sàng và điều trị, Tạp chí Truyền nhiễm Việt Nam, Vol. 2, No. 50, pp. 131-138, 2025. DOI: 10.59873/vjid.v2i50.476
- M. H. Norris et al., Expanding the molecular epidemiology of melioidosis in North Central Vietnam, Plos Neglected Tropical Diseases, Vol. 20, No. 2, p. e0013945, 2026. DOI: 10.1371/journal.pntd.0013945
- I. Gassiep et al., Laboratory diagnosis of melioidosis, Plos Neglected Tropical Diseases, Vol. 19, No. 12, p.e0013761, 2025. DOI: 10.1371/journal.pntd.0013761
- K. Selvam , M. A. Najib, M. F. Khalid, A. Harun and I. Aziah, Performance of antibody-detection tests for human melioidosis: A systematic review and meta-analysis, Malaysian Journal of Medical Sciences, Vol. 31, No. 6, pp. 34-56, 2024. DOI: 10.21315/mjms2024.31.6.4
- G. E. Wagner et al., Melioidosis DS rapid test: A standardized serological dipstick assay with increased sensitivity and reliability due to multiplex detection, Plos Neglected Tropical Diseases, Vol. 14, No. 8, p. e0008452, 2020. DOI: 10.1371/journal.pntd.0008452
- P. Amornchai et al., Evaluation of antigen-detecting and antibody-detecting diagnostic test combinations for diagnosing melioidosis, Plos Neglected Tropical Diseases, Vol. 15, No. 11, p. e0009840, 2021. DOI: 10.1371/journal.pntd.0009840
- Q. T. L. Tran et al., Clinical utility of combined whole-cell antigen and recombinant hemolysis co-regulated protein 1-enzyme-linked immunosorbent assays reveals underdiagnosed cases of melioidosis in Vietnam, American Journal of Tropical Medicine and Hygiene, Vol. 107, No. 3, pp. 585–91, 2022. DOI: 10.4269/ajtmh.21-1143
- P. Wu et al., Anti-Hcp1 monoclonal antibody is protective against Burkholderia pseudomallei via recognizing amino acids at Asp95-Leu114, Pathogens, Vol. 13, No. 1, p. 43, 2024. DOI: 10.3390/pathogens13010043
- B. Zhang et al., Molecular mechanisms of AhpC in resistance to oxidative stress in Burkholderia thailandensis, Frontiers in Microbiology, Vol.10, p. 1483, 2019. DOI: 10.3389/fmicb.2019.01483
- K. Selvam, M. A. Najib, M. F. Khalid, A. Harun and I. Aziah, Performance of antibody-detection tests for human melioidosis: A systematic review and meta-analysis, Malaysian Journal of Medical Sciences, Vol. 31, No. 6, p. 34-56, 2024. DOI: 10.21315/mjms2024.31.6.4
- F. Du et al., Regulating the T7 RNA polymerase expression in E. coli BL21 (DE3) to provide more host options for recombinant protein production, Microbial Cell Factories, Vol. 20, No. 1, p. 189, 2021. DOI: 10.1186/s12934-021-01680-6
- A. R. Tuttle, N. D. Trahan and M. S. Son, Growth and maintenance of Escherichia coli laboratory strains, Current Protocols, Vol. 1, No. 1, p. e20, 2021. DOI: 10.1002/cpz1.20
- Nandi T et al., Burkholderia pseudomallei sequencing identifies genomic clades with distinct recombination, accessory, and epigenetic profiles, Genome Research, Vol. 25, No. 1, pp. 129-41, 2015. DOI: 10.1101/gr.177543.114
- T. T Hong et al., Expression and characterization of a new serine protease inhibitory protein in Escherichia coli, Biomedical Research and Therapy, Vol. 7, No. 2, pp. 3633-3644, 2020. DOI: 10.15419/bmrat.v7i2.590
- Trịnh Văn Toàn, Lê Ngọc Diệp, Hoàng Đăng Hiếu, Phạm Việt Hùng, Võ Viết Cường và Lê Thị Lan Anh, Khảo sát điều kiện nuôi cấy biểu hiện chuỗi nhẹ độc tố thần kinh type huyết thanh B tái tổ hợp của vi khuẩn Clostridium botulinum trong tế bào Escherichia coli BL21 (DE3), Tạp chí Khoa học và Công nghệ nhiệt đới, số 35, 2024. DOI: 10.58334/vrtc.jtst.n35.06
- Trần Thị Lệ Quyên et al., Đánh giá kháng nguyên tái tổ hợp hemolysin co-regulated protein 1 (Hcp1) trong chẩn đoán nhanh bệnh nhân nhiễm melioidosis (bệnh Whitmore) bằng kỹ thuật ELISA, Bản B của Tạp Chí Khoa học và Công nghệ Việt Nam, Vol. 62, No. 9, 2020.