Đánh giá tình trạng kháng kháng sinh của một số vi khuẩn phân lập từ cá chép Cyprinidae tại Cần Thơ và An Giang
Viện Y sinh Nhiệt đới, Trung tâm Nhiệt đới Việt - Nga, Hà Nội, Việt Nam
63 Nguyễn Văn Huyên, Nghĩa Đô, Hà Nội
Email: pvnhung0506@gmail.com
Nội dung chính của bài viết
Tóm tắt
Nghiên cứu này cung cấp dữ liệu thực tế về tình trạng kháng kháng sinh của các chủng vi khuẩn phân lập từ cá chép tại tỉnh Cần Thơ, An Giang, Việt Nam. Tổng cộng 200 mẫu (vẩy, da, mô cơ và các cơ quan nội tạng) từ cá chép được thu thập từ các ao hồ tự nhiên và một số cơ sở nuôi trồng thủy sản tại Cần Thơ, An Giang. Các mẫu được tiến hành làm sạch, khử trùng sau đó tiến hành đồng nhất và nuôi cấy phân lập trên các môi trường khác nhau. Các chủng vi khuẩn được định danh bằng hệ thống MALDI-TOF MS và đánh giá độ nhạy cảm với kháng sinh bằng phương pháp khuếch tán đĩa Kirby-Bauer. Kết quả chúng tôi đã phân lập được 607 chủng vi khuẩn đồng thời ghi nhận kiểu hình kháng kháng sinh ở nhiều nhóm vi khuẩn. Cụ thể, các chủng thuộc chi Enterobacter và Serratia có tỉ lệ kháng lên tới 90,6–100% với amoxicillin/acid clavulanic. Trong khi đó, Hafnia alvei thể hiện tỷ lệ kháng cao với cephalosporin thế hệ 3 (81,3%) và kháng trung bình với cephalosporin thế hệ 4 (25%). Ở vi khuẩn Pseudomonas spp. cho thấy sự đa kháng, các chủng vi khuẩn này thể hiện tính kháng 100% với ciprofloxacin, cephalosporin (thế hệ 3 và 4) và imipenem. Tương tự, toàn bộ chủng Acinetobacter được phân lập đều kháng ciprofloxacin (100%). Đối với chi Aeromonas, tỷ lệ kháng levofloxacin là 61,8%; đồng thời 34,7% chủng kháng với ít nhất một kháng sinh khác trong nhóm fluoroquinolone. Những phát hiện này phản ánh thực trạng kháng kháng sinh đang tồn tại ở các vi khuẩn phân lập từ động vật thủy sản cho thấy sự cần thiết của việc tiếp tục theo dõi và đánh giá tình trạng kháng kháng sinh trong lĩnh vực thủy sản.
Abstract
Assessment of antibiotic resistance in bacteria isolated from Cyprinidae fish in Can Tho and An Giang
This study provided empirical data on the antibiotic resistance profiles of bacteria isolated from fish of the family Cyprinidae in Can Tho and An Giang provinces, Vietnam. A total of 200 samples including scales, skin, muscle tissue, and internal organs were collected from Cyprinidae obtained from natural ponds, lakes, and aquaculture facilities. The samples were cleaned, surface-sterilized, homogenized, and cultured for bacterial isolation on different media. Bacterial isolates were identified using the MALDI-TOF MS system, and antibiotic susceptibility was evaluated using the Kirby-Bauer disk diffusion method. The results showed that 607 bacterial strains were isolated, and alarming antibiotic resistance phenotypes were recorded in multiple bacterial groups. Specifically, isolates from the genera Enterobacter and Serratia exhibited extremely high resistance rates (90.6–100%) to amoxicillin/clavulanic acid. Meanwhile, Hafnia alvei showed high resistance to third-generation cephalosporins (81.3%) and moderate resistance to fourth-generation cephalosporins (25%). Pseudomonas spp. demonstrated multidrug resistance, with all isolates exhibiting 100% resistance to ciprofloxacin, third- and fourth-generation cephalosporins, and imipenem. Similarly, all recovered Acinetobacter isolates were resistant to ciprofloxacin (100%). Among Aeromonas isolates, the resistance rate to levofloxacin reached 61.8%, and 34.7% were resistant to at least one additional fluoroquinolone. These findings reflect the existing status of antibiotic resistance among bacterial isolates from aquatic animals and highlight the need for continued surveillance and assessment of antibiotic resistance in aquaculture.
Key words: Antibiotic-resistant; Aeromonas spp.; Enterobacteriaceae; Enterococcus spp.; resistance rates.
Từ khóa
Aeromonas spp., Enterobacteriaceae, Enterococcus spp., kháng kháng sinh, tỉ lệ kháng
Chi tiết bài viết

Bài báo này được cấp phép theo Creative Commons Attribution 4.0 International License.
Điểm nổi bật
Nghiên cứu đã phân lập được tổng cộng 607 chủng vi khuẩn gồm 53 loài thuộc 20 chi và 12 họ từ 200 mẫu cá chép thu thập tại một số sông, hồ tự nhiên và cơ sở nuôi trồng thủy sản thuộc tỉnh Cần Thơ và An Giang. Vi khuẩn Gram âm chiếm ưu thế (với 431 chủng/19 chi), cao nhất là trực khuẩn kỵ khí tùy ý (64,7%). Trong khi vi khuẩn Gram dương chỉ gồm một chi Enterococcus nhưng chiếm tỷ lệ cao (29,0%).
Nghiên cứu cho thấy tình trạng kháng kháng sinh phổ biến ở các chủng vi khuẩn phân lập, đặc biệt là kháng các nhóm cephalosporin thế hệ mới và fluoroquinolone. Những phát hiện này cho thấy sự tồn tại của vi khuẩn kháng thuốc trong môi trường thủy sinh được khảo sát, qua đó nhấn mạnh tầm quan trọng của việc tiếp tục theo dõi và đánh giá tình trạng kháng kháng sinh trong lĩnh vực thủy sản.
1. ĐẶT VẤN ĐỀ
Kháng kháng sinh hiện được ghi nhận rộng rãi trong nhiều hệ sinh thái khác nhau trên thế giới. Trong lĩnh vực nông nghiệp, chăn nuôi và đặc biệt là nuôi trồng thủy sản, kháng sinh được sử dụng nhằm kiểm soát dịch bệnh, và việc sử dụng này được cho là có liên quan đến sự xuất hiện của các chủng vi khuẩn kháng thuốc trong môi trường nước [1,2]. Một số nghiên cứu trước đây cho thấy việc sử dụng kháng sinh kéo dài có thể liên quan đến sự gia tăng tỷ lệ vi khuẩn mang đặc điểm kháng thuốc trong các hệ sinh thái tự nhiên, qua đó phản ánh mức độ tác động của hoạt động con người đối với môi trường [3,4].
Các loài thủy sinh như cá và ốc đã được ghi nhận có thể mang vi khuẩn kháng kháng sinh trong cơ thể, từ đó có khả năng góp phần vào sự phân bố của các chủng này trong môi trường nước [4,5]. Bên cạnh đó, hiện tượng chuyển gen ngang giữa các loài vi khuẩn đã được mô tả trong nhiều môi trường khác nhau, bao gồm cả môi trường bệnh viện, nơi thường quan sát thấy sự đồng tồn tại của gen kháng thuốc và gen độc lực [6]. Một số nghiên cứu cho thấy trong điều kiện có dư lượng kháng sinh, quá trình trao đổi vật chất di truyền giữa các vi khuẩn có thể xảy ra, song vai trò cụ thể của quá trình này trong hệ sinh thái thủy sinh chưa được làm sáng tỏ đầy đủ.
Nhiều nhóm vi khuẩn khác nhau đã được phân lập từ đường ruột cá nuôi, trong số đó có các chủng biểu hiện đặc tính kháng kháng sinh. Các chi vi khuẩn thường được phân lập từ cá bao gồm Acinetobacter, Aeromonas, Citrobacter, Enterococcus, Escherichia, Morganella, Plesiomonas, Proteus và Shewanella [7–9]. Trong số này, Aeromonas, Enterococcus, Escherichia, Plesiomonas và Proteus được ghi nhận với tần suất tương đối cao. Việc phát hiện Escherichia coli, Enterococcus spp. và Proteus spp. trong môi trường nước và sinh vật thủy sinh thường được sử dụng như các chỉ thị về ô nhiễm phân người và động vật [10,11]. Đồng thời, một số nghiên cứu cũng cho thấy các nhóm vi khuẩn này có thể mang gen kháng thuốc dễ dàng lan truyền.
Các nghiên cứu trước đây ghi nhận rằng một số chủng vi khuẩn phân lập từ cá có thể mang đồng thời gen kháng thuốc và gen liên quan đến độc lực. Ví dụ, Enterococcus spp. được mô tả có thể mang gen kháng vancomycin và có khả năng trao đổi vật chất di truyền thông qua plasmid hoặc transposon [12]. Tương tự, Aeromonas spp. cũng đã được ghi nhận mang nhiều yếu tố liên quan đến độc lực [13]. Điều này cho thấy môi trường thủy sinh và cá nuôi có thể đóng vai trò như “ổ chứa” quan trọng của các tác nhân kháng thuốc, không chỉ lưu giữ mà còn thúc đẩy sự lan truyền của các tác nhân kháng thuốc ra môi trường [14].
Sông Mekong là hệ thống sông lớn của khu vực Đông Nam Á, đóng vai trò quan trọng trong khai thác và nuôi trồng thủy sản, góp phần đáng kể vào phát triển kinh tế và an ninh lương thực của Việt Nam [15]. Cá nước ngọt là nhóm sinh vật tiếp xúc trực tiếp với môi trường nước và có thể phản ánh đặc điểm vi sinh vật của hệ sinh thái này. Một số nghiên cứu đã ghi nhận cá nước ngọt mang các chủng vi khuẩn kháng với ít nhất một loại kháng sinh được thử nghiệm, trong đó có các chi như Aeromonas, Pseudomonas và Acinetobacter. Tỷ lệ kháng tương đối cao đã được quan sát thấy đối với Amoxicillin-Clavulanic acid, Cefepime, Cefoxitin, Aztreonam và Ceftazidime trong một số khảo sát [16].
Trên cơ sở đó, nghiên cứu này khảo sát đặc điểm kháng kháng sinh của các chủng vi khuẩn phân lập từ cá thuộc họ Cyprinidae thu tại một số địa điểm ở tỉnh Cần Thơ và An Giang, nhằm cung cấp thêm dữ liệu thực nghiệm cho lĩnh vực vi sinh môi trường.
2. VẬT LIỆU VÀ PHƯƠNG PHÁP
2.1. Vật liệu
Đối tượng nghiên cứu: Tổng cộng 200 mẫu cá thuộc họ Cyprinidae ở độ tuổi từ 1 đến 3 năm, được đánh bắt bằng lưới hoặc câu trong khoảng thời gian từ tháng 4 đến tháng 5 năm 2025 tại một số sông, hồ tự nhiên và cơ sở nuôi trồng thủy sản thuộc tỉnh Cần Thơ và An Giang. Mỗi mẫu được thu từ một cá thể riêng biệt, bao gồm vẩy, da, mô cơ và các cơ quan nội tạng. Sau đó được vận chuyển lạnh về Phòng thí nghiệm của Trung tâm Nhiệt đới Việt - Nga tại Tp. Hồ Chí Minh để nghiên cứu. Từ các mẫu này 607 chủng vi khuẩn đã được phân lập và định danh.
Môi trường sử dụng nuôi cấy và phân lập vi khuẩn:
- Môi trường thạch Endo (HiMedia, Ấn Độ, M029);
- Môi trường Enterococci agar (HiMedia, Ấn Độ);
- Môi trường Xylose Lysine Deoxycholate (XLD) agar (HiMedia, Ấn Độ, M031);
- Môi trường thạch dinh dưỡng (Nutrient agar) (HiMedia, Ấn Độ).
Các nhóm vi khuẩn được thử nghiệm với các loại kháng sinh theo Clinical and Laboratory Standards Institute (CLSI, 2023), đặc điểm sinh học - cơ chế kháng thuốc của từng nhóm vi khuẩn cụ thể trong Bảng 1:
Bảng 1. Các loại kháng sinh được sử dụng trong kiểm tra tính nhạy cảm theo nhóm vi khuẩn
Nhóm vi khuẩn | Kháng sinh thử nghiệm |
Enterobacteriaceae | ciprofloxacin, amikacin, ceftazidime, cefotaxime, cefepime, amoxicillin/acid clavulanic, imipenem, meropenem |
Vi khuẩn Gram âm không lên men | ciprofloxacin, amikacin, ceftazidime, cefepime, imipenem, meropenem |
Aeromonas spp. | ciprofloxacin, levofloxacin, ceftazidime, cefepime, cotrimoxazole |
Enterococcus spp. | ciprofloxacin, gentamicin, vancomycin, ampicillin, norfloxacin |
2.2. Phương pháp nghiên cứu
2.2.1. Phân lập và định danh vi khuẩn
Trước khi xử lý, bề mặt mẫu được làm sạch và khử trùng theo quy trình vô trùng, sau đó các phần mô được lấy trong điều kiện vô trùng và đồng nhất trong dung dịch nước muối sinh lý 0,9% (NaCl) với tỷ lệ 1:9 bằng thiết bị đồng nhất mẫu.
Dịch đồng nhất được cấy trực tiếp trên các môi trường thạch Endo để phân lập các vi khuẩn Gram âm đường ruột; môi trường XLD agar để phát hiện và phân lập Salmonella spp. cũng như một số vi khuẩn gây bệnh khác (ủ ở 37°C trong 24 giờ) và môi trường Enterococci agar để phân lập vi khuẩn thuộc chi Enterococcus spp. (ủ ở 37°C trong 48 giờ). Sau thời gian ủ, tiến hành quan sát hình thái của khuẩn lạc trên các đĩa môi trường, lựa chọn khuẩn lạc dựa trên đặc điểm hình thái đặc trưng và cấy truyền trên môi trường thạch dinh dưỡng nhằm thu nhận các chủng vi khuẩn thuần khiết [17].
Các chủng vi khuẩn thuần được tiến hành định danh bằng hệ thống MALDI-TOF MS (Bruker, Đức) sử dụng bộ kit IVD Matrix HCCA-portioned (Bruker, Đức) theo hướng dẫn của nhà sản xuất, kết quả được xác nhận ở mức loài khi điểm số (score) ≥ 2 đảm bảo độ tin cậy cao; nếu điểm số thấp hơn, vi khuẩn chỉ được định danh tới cấp chi.
2.2.2. Thử nghiệm kháng kháng sinh
Khả năng kháng kháng sinh của các chủng vi khuẩn được xác định bằng phương pháp khuếch tán đĩa Kirby-Bauer trên môi trường thạch Mueller-Hinton (Himedia, Ấn Độ), khoanh giấy kháng sinh (Oxoid, Mỹ) theo tài liệu hướng dẫn của Viện Tiêu chuẩn Lâm sàng và Phòng thí nghiệm Hoa Kỳ [18]. Vi khuẩn sinh β-lactamase phổ rộng (ESBL) được phát hiện bằng kỹ thuật khuếch tán đĩa đôi trên môi trường thạch Mueller-Hinton [19]. Chủng vi khuẩn Escherichia coli ATCC 25922 và chủng Enterococcus faecalis ATCC 29212 được sử dụng cho tất cả các thí nghiệm làm đối chứng dương.
Kết quả đánh giá dựa trên đường kính vòng ức chế (mm) và phân loại mức độ nhạy cảm theo tiêu chuẩn CLSI M100 (năm 2023). Các chủng vi khuẩn được xác định là đa kháng kháng sinh (multi-drug resistant, MDR) khi có kiểu hình kháng đối với ít nhất một kháng sinh ở ≥ 3 nhóm kháng sinh khác nhau [20].
2.2.3. Phân tích thống kê
Dữ liệu được xử lý và phân tích bằng Microsoft Excel. Tỷ lệ kháng kháng sinh của các chủng vi khuẩn được tính theo phần trăm (%), mỗi chủng được coi là một đơn vị quan sát độc lập.
3. KẾT QUẢ
3.1. Kết quả phân lập các chủng vi khuẩn trên mẫu cá
Kết quả phân lập vi khuẩn từ mẫu cá được trình bày trong Bảng 2. Trực khuẩn Gram âm kỵ khí tùy ý gồm 393 chủng (64,2%), vi khuẩn Gram âm không lên men gồm 32 chủng (5,8%) và vi khuẩn Gram âm hiếu khí gồm 6 chủng (1,0%). Cầu khuẩn Gram dương bao gồm vi khuẩn thuộc chi Enterococcus, gồm 176 chủng (29,0%).
Các chủng vi khuẩn phân lập thuộc 12 họ: Aeromonadaceae, Morganellaceae, Enterobacteriaceae, Hafniaceae, Shewanellaceae, Yersiniaceae, Moraxellaceae, Pseudomonadaceae, Flavobacteriaceae, Vibrionaceae, Alcaligenaceae và Enterococcaceae. Phân tích chi tiết cho thấy sự hiện diện của nhiều loại vi khuẩn gram âm trong các mẫu cá chép được nghiên cứu bao gồm: Aeromonas (A. veronii, A. hydrophila, A. ichtniosmia, A. caviae, A. salmonicida, A. jandaei, A. enteropelogenes, A. bestiarum), Proteus (P. mirabilis, P. Vulgaris, P. hauseri, P. penneri), Klebsiella (K. pneumoniae, K. variicola, K. aerogenes, K. oxytoca), Enterobacter (E. cloacae, E. kobei, E. asburiae, E. hormaechei), Citrobacter (C. freundii, C. braakii), Serratia (S. fonticola, S. marcescens), Snewanella (S. algae, S. xiamenensis), Providencia (P. alcalifaciens, P. rettgeri), và một số chủng riêng lẻ được phân lập Alcaligenes faecalis, Hafnia alvei, Moellerella wisconsis, Morganella morganii, Myroides injenensis, Plesiomonas shigelloides, Raoultella ornithinolytica và Vibrio tasmaniensis.
Bảng 2. Cấu trúc các nhóm vi khuẩn phân lập từ 200 mẫu cá chép Cyprinidae.
Nhóm vi khuẩn | Số lượng | Tỷ lệ (%) | ||
| Họ | Chi | ||
Vi khuẩn gram âm kỵ khí tùy ý | Aeromonadaceae | Aeromonas spp. | 128 | 21,09 |
Morganellaceae | Proteus spp. | 87 | 14,33 | |
Providencia spp. | 19 | 3,13 | ||
Morganella morganii | 18 | 2,97 | ||
Moellerella wisconsis | 4 | 0,66 | ||
Enterobacteriaceae | Klebsiella spp. | 27 | 4,45 | |
Enterobacter spp. | 22 | 3,62 | ||
Escherichia coli | 21 | 3,46 | ||
Citrobacter spp. | 16 | 2,64 | ||
Plesiomonas shigelloides | 8 | 1,32 | ||
Raoultella ornithinolytica | 13 | 2,14 | ||
Hafniaceae | Hafnia alvei | 16 | 2,64 | |
Shewanellaceae | Snewanella spp. | 12 | 1,98 | |
Yersiniaceae | Serratia spp. | 2 | 0,33 | |
Vi khuẩn Gram âm không lên men | Moraxellaceae | Acinetobacter spp. | 7 | 1,15 |
Pseudomonadaceae | Pseudomonas spp. | 25 | 4,12 | |
Vi khuẩn Gram âm hiếu khí | Flavobacteriaceae | Myroides injenensis | 2 | 0,33 |
Vibrionaceae | Vibrio spp. | 3 | 0,49 | |
Alcaligenaceae | Alcaligenes faecalis | 1 | 0,16 | |
Cầu khuẩn Gram dương | Enterococcaceae | Enterococcus spp. | 176 | 29,00 |
Tổng |
|
| 607 | 100 |
Bên cạnh các vi khuẩn Gram âm đường ruột, nghiên cứu cũng phân lập được nhóm vi khuẩn Gram âm không lên men, chủ yếu thuộc các chi Pseudomonas (P. aeruginosa, P. otitidis, P. putida, P. monteili, P. quariconensis) và Acinetobacter (A. baumannii, A. junii, A. lactucae).
Với nhóm cầu khuẩn Gram dương, Enterococcus là chi chiếm ưu thế tuyệt đối. Thành phần loài của chi này rất đa dạng, với sự hiện diện của E. faecalis, E. faecium, E. hirae, E. casseliflavus, E. pseudoavium, E. thailandicus, E. durans, E. gallinarum, và E. raffinosus. Trong đó, hai loài E. faecalis (51,14%) và E. faecium (33,52%) là phổ biến nhất, cùng chiếm tỷ lệ áp đảo.
3.2. Đặc điểm kháng kháng sinh của các chủng vi khuẩn phân lập
Đặc điểm kháng kháng sinh của vi khuẩn gram âm phân lập từ cá cho thấy có sự khác biệt rõ rệt về khả năng kháng amoxicillin/acid clavulanic ở các chủng. Trong khi các nhóm vi khuẩn Serratia spp. (100%), Enterobacter spp. (90,9%), M. morganii (87,7%), Citrobacter spp. (85,7%) và H. alvei (87,5%) có tỉ lệ kháng rất cao, thì nhóm Providencia spp. (36,8%), Proteus spp. (34,5%), Klebsiella spp. (33,3%) và Escherichia coli (26,7%) lại có tỉ lệ kháng thấp hơn hẳn chỉ chiếm một phần ba tổng số các chủng được phân lập (Hình 1a).
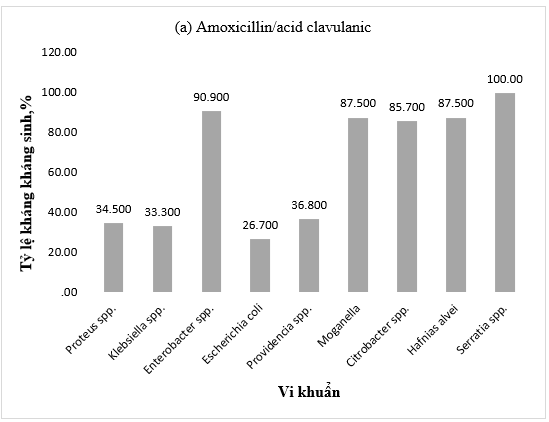
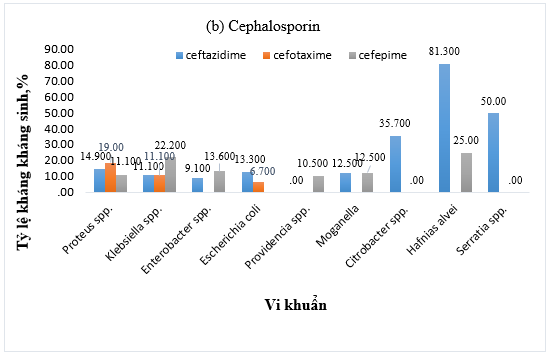
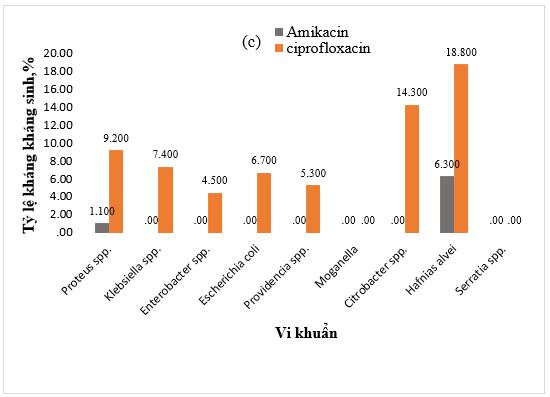
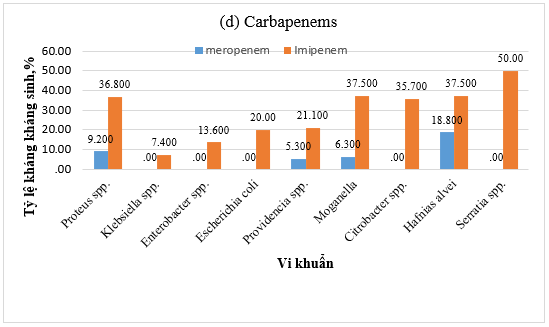
Hình 1. Tỷ lệ kháng kháng sinh (a) Amoxicillin/acid clavulanic, (b) Cephalosporin, (c) Amikacin và ciprofloxacin, (d) Carbapenem của vi khuẩn Gram âm
Tỷ lệ kháng kháng sinh nhóm cephalosporin thế hệ 3 biến đổi đáng kể giữa các chủng vi khuẩn. Trong khi hầu hết các chủng Proteus spp., E. coli, M. morganii, Klebsiella spp., Enterobacter spp. và Providencia spp. có tỷ lệ kháng thấp (dưới 14,9%), thì tỷ lệ này lại cao hơn ở Citrobacter spp. và Serratia spp. (50,0%), và đặc biệt cao ở H. Alvei (81,3%). Ngược lại, tất cả các chủng đều thể hiện độ nhạy cảm cao hơn đối với cephalosporin thế hệ 4, với tỷ lệ kháng được ghi nhận dưới 25,0% (Hình 1b).
Phần lớn các chủng vi khuẩn Gram âm phân lập được đều nhạy cảm cao (100%) đối với nhóm aminoglycoside, ngoại trừ các chủng H. alvei và Proteus spp. Đối với kháng sinh ciprofloxacin, tỷ lệ kháng quan sát được trong số các chủng còn lại là tương đối thấp (<18,8%) (Hình 1c).
Về nhóm kháng sinh carbapenem, tỷ lệ kháng imipenem ở Serratia spp. được ghi nhận là dưới 50%. Trong khi đó, tỷ lệ kháng meropenem thấp hơn đáng kể: dưới 37,5% ở các chủng M. morganii, H. alvei, Citrobacter spp. và Proteus spp., và dưới 18,8% ở các chủng khác (Hình 1d).
Đáng chú ý, các chủng sinh enzyme β-lactamase phổ rộng (ESBL), nguyên nhân chính gây kháng các cephalosporin thế hệ 3 và penicillin đã được phát hiện với tỷ lệ khác nhau giữa các chi vi khuẩn: cao nhất ở Hafnia alvei (6,3%), tiếp theo là Klebsiella spp. (3,7%) và Proteus spp. (1,1%) (Hình 2).
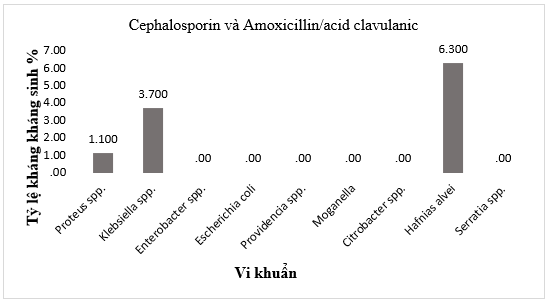
Hình 2. Các chủng vi khuẩn Gram âm sinh β-lactamase phổ rộng
Với 128 chủng Aeromonas spp. được phân lập chiếm tỉ lệ (21,1%), kết quả định danh xác định được 8 loài, trong đó Aeromonas veronii (41,41%) và Aeromonas hydrophila (16,41%) là những loài chiếm ưu thế. Đánh giá đặc điểm kháng kháng sinh cho thấy các chủng Aeromonas spp. kháng levofloxacin ở mức cao (61,8%) và kháng fluoroquinolone ở mức trung bình (34,7%) (Hình 3).
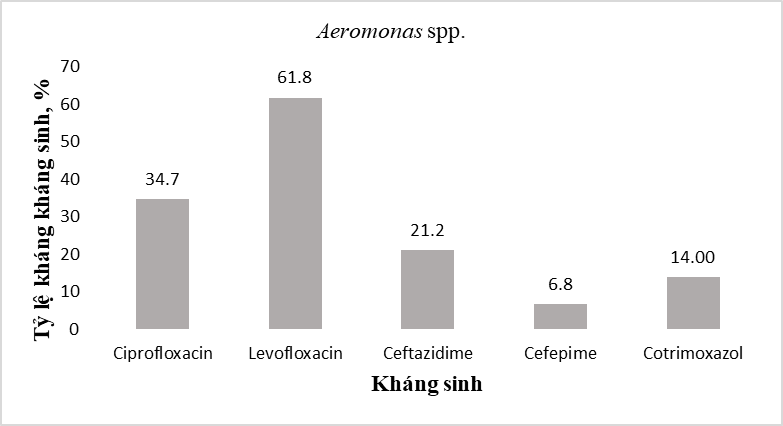
Hình 3. Tỷ lệ kháng kháng sinh của các chủng Aeromonas spp.
Kết quả phân tích khả năng kháng kháng sinh với các vi khuẩn thuộc chi Acinetobacter cho thấy tỷ lệ kháng (100%) với ciprofloxacin. Trái ngược với điều này, tất cả các chủng Acinetobacter đều cho thấy độ nhạy cảm hoàn toàn (100%) đối với amikacin, carbapenem (imipenem, meropenem) và cotrimoxazol. Trong khi đó, ở chi Pseudomonas, tình trạng kháng kháng sinh được ghi nhận phức tạp hơn khi các chủng này kháng hoàn toàn (100%) đối với ciprofloxacin, cephalosporin thế hệ 3, 4 và imipenem. Dù vậy, tỷ lệ kháng meropenem thấp hơn rõ rệt (16,7%) và amikacin vẫn duy trì hiệu quả 100% trên toàn bộ chủng nghiên cứu (Hình 4).
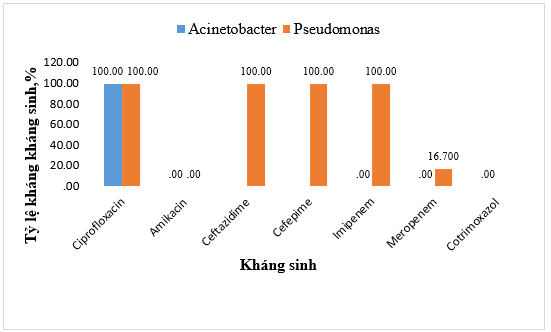
Hình 4. Tỷ lệ kháng kháng sinh của vi khuẩn Pseudomonas và Acinetobacter
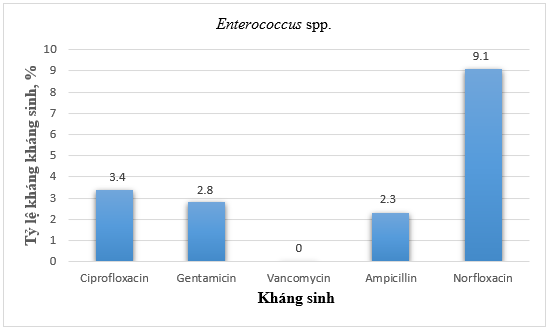
Hình 5. Tỷ lệ kháng kháng sinh của vi khuẩn Enterococcus spp.
Khi phân tích đặc điểm kháng kháng sinh của vi khuẩn Enterococcus spp. cho thấy, tất cả các chủng đều nhạy cảm với vancomycin (Hình 5). Tỷ lệ các chủng kháng ciprofloxacin, gentamicin và ampicillin không quá 3,4%, tỷ lệ kháng norfloxacin 9,1%.
4. THẢO LUẬN
Kết quả nghiên cứu cho thấy các chủng vi khuẩn phân lập trên cá có độ đa dạng cao, bao gồm cả vi khuẩn Gram âm và Gram dương. Trong đó, các trực khuẩn Gram âm kỵ khí tùy ý chiếm ưu thế (64,2%), phản ánh đúng đặc điểm sinh thái giàu chất hữu cơ của môi trường nuôi cá nước ngọt, nơi các họ Aeromonadaceae, Enterobacteriaceae và Vibrionaceae thường phát triển mạnh. Đáng chú ý, chi Aeromonas (A. veronii, A. hydrophila, A. caviae, A. salmonicida) xuất hiện với tần suất cao, khẳng định vai trò là nhóm vi khuẩn bản địa, có tiềm năng gây bệnh cơ hội quan trọng ở cá nước ngọt [21,22].
Việc không phát hiện các vi khuẩn chỉ thị của sự ô nhiễm phân cấp tính Salmonella và Shigella cho thấy tại thời điểm nghiên cứu không có sự ô nhiễm nguồn phân mới với tải lượng cao vào môi trường nước. Tuy nhiên, sự hiện diện của các chi Proteus, Klebsiella, Enterobacter, Citrobacter, Serratia và Morganella vốn là những chỉ thị đặc trưng cho đường ruột của người và động vật máu nóng lại cho thấy tình trạng ô nhiễm phân đã tồn tại và kéo dài trong hệ thống nuôi trồng thủy sản [23].
Nhóm vi khuẩn Gram âm không lên men chiếm tỷ lệ thấp (5,3%), tuy nhiên nhóm này bao gồm nhiều loài có độc lực và khả năng kháng kháng sinh đáng kể, như Pseudomonas và Acinetobacter [24,25]. Trong khi đó nhóm vi khuẩn Gram dương, Enterococcus chiếm tỉ lệ (29,0%), chủ yếu là E. faecalis và E. faecium, được xem là chỉ thị quan trọng phản ánh điều kiện vệ sinh môi trường và nguy cơ lan truyền gen kháng kháng sinh [11,26]. Tổng cộng, các chủng phân lập được thuộc 12 họ và hơn 20 chi, phù hợp với xu hướng được báo cáo trong nhiều nghiên cứu gần đây tại Việt Nam và khu vực, khi Aeromonas, Enterobacteriaceae và Enterococcus là ba nhóm phổ biến nhất [8]. Sự xuất hiện của một số vi khuẩn môi trường khác như Shewanella, Vibrio, Flavobacterium và Alcaligenes càng chứng tỏ mối liên hệ mật thiết giữa điều kiện nuôi trồng và sự phong phú của các chủng vi khuẩn phân lập được [27,28].
Kết quả phân tích cho thấy sự khác biệt đáng kể về mức độ kháng kháng sinh giữa các chi vi khuẩn Gram âm. Trong số 128 chủng Aeromonas phân lập, hai loài chiếm ưu thế là A. veronii (41,41%) và A. hydrophila (16,41%) đều thể hiện tỷ lệ kháng levofloxacin cao (61,8%) và kháng fluoroquinolone ở mức trung bình (34,7%). Khác với Aeromonas, Acinetobacter spp. cho thấy phổ kháng hẹp hơn, chỉ kháng mạnh với ciprofloxacin trong khi vẫn nhạy cảm hoàn toàn với amikacin, carbapenem (imipenem, meropenem) và cotrimoxazol. Đáng chú ý là Pseudomonas spp. mặc dù vẫn còn nhạy cảm với amikacin nhưng lại cho thấy tính kháng rất cao (100%) đối với nhiều nhóm kháng sinh then chốt: ciprofloxacin, cephalosporin thế hệ 3 và 4, và cả imipenem. Kiểu hình đa kháng (MDR) này ở Pseudomonas được cho là liên quan đến các cơ chế phức tạp như hệ thống bơm đẩy đa kháng và giảm tính thấm của màng ngoài tế bào [29]. Tuy nhiên, cơ chế cụ thể trong các chủng của nghiên cứu hiện tại chưa được xác định.
Họ Enterobacteriaceae bao gồm các chi Serratia, Enterobacter, Morganella, Citrobacter và Hafnia alvei thể hiện tỷ lệ kháng cao với amoxicillin/acid clavulanic (≥ 85%), phù hợp với khả năng sản xuất β-lactamase của nhóm này [30,31]. Trong khi đó, tỷ lệ kháng với cephalosporin thế hệ 3 (C3G) biến động lớn: thấp (< 15%) ở Proteus, Escherichia coli, Morganella, Klebsiella, Enterobacter, Providencia; nhưng lại rất cao ở Citrobacter, Serratia và đặc biệt là H. alvei (81,3%). Cephalosporin thế hệ 4 (C4G) vẫn duy trì hiệu lực tương đối (tỷ lệ kháng ≤25%), song xu hướng kháng tăng dần đã được ghi nhận tương tự tại Việt Nam [32].
Các nhóm khác: Aminoglycoside (như amikacin, gentamicin) vẫn cho hiệu quả tốt trên hầu hết các chủng, ngoại trừ H. alvei và Proteus. Tỷ lệ kháng ciprofloxacin ở mức thấp (<18,8%) ở phần lớn các chi, dù đã xuất hiện một số dấu hiệu đề kháng. Carbapenem (imipenem, meropenem) tiếp tục thể hiện phổ hiệu lực rộng, ngoại trừ Serratia spp. (tỷ lệ kháng imipenem lên tới 50%). Các chủng sinh enzym kháng β-lactam phổ rộng được ghi nhận ở Proteus (1,1%), Klebsiella (3,7%) và H. alvei (6,3%), đặc điểm kiểu hình này có thể liên quan đến cơ chế sinh β-lactamase phổ rộng (ESBL) phù hợp với các báo cáo trước đây về việc ESBL được ghi nhận ngày càng nhiều trong môi trường thủy sản [33,34].
Đối với nhóm Gram dương, tất cả các chủng Enterococcus phân lập được vẫn nhạy cảm với vancomycin. Tỷ lệ kháng với ciprofloxacin, gentamicin và ampicillin còn ở mức thấp (≤ 3,6%). Tuy nhiên, việc giám sát liên tục là cần thiết do vai trò trung gian quan trọng của Enterococcus trong việc lan truyền gen kháng kháng sinh.
Hạn chế của nghiên cứu: Nghiên cứu này vẫn còn một số hạn chế. Phạm vi khảo sát chỉ giới hạn ở các vi khuẩn phân lập từ mẫu cá thu thập tại Cần Thơ và An Giang, và chưa thực hiện phân tích so sánh giữa các nhóm vi khuẩn phân lập từ những môi trường sống khác nhau (cá nuôi và cá tự nhiên). Bên cạnh đó, phương pháp nuôi cấy chọn lọc có thể chưa phản ánh đầy đủ tính đa dạng của quần thể vi khuẩn hiện diện trong mẫu. Đơn vị phân tích của nghiên cứu là các chủng phân lập thay vì cá thể vật chủ, do đó chưa đánh giá được mức độ phân bố đặc tính kháng ở cấp độ quần thể cá. Ngoài ra, nghiên cứu chưa tiến hành xác định cơ chế kháng ở mức gen. Vì vậy, các kết quả thu được chủ yếu phản ánh đặc điểm kháng kháng sinh của các chủng vi khuẩn được phân lập trong phạm vi khảo sát. Các kết quả mang tính khảo sát ban đầu và cần được kiểm chứng trong các nghiên cứu tiếp theo.
5. KẾT LUẬN
Nghiên cứu đã phân lập được tổng cộng 607 chủng vi khuẩn, thể hiện sự đa dạng sinh học đáng kể với 53 loài thuộc 20 chi và 12 họ. Trong đó, nhóm vi khuẩn Gram âm thể hiện qua tính đa dạng cao (11 họ, 19 chi), trực khuẩn kỵ khí tùy tiện là nhóm chiếm đa số với 393 chủng (64,7%). Trong khi đó, mặc dù kém đa dạng hơn (chỉ một chi), chi Enterococcus ở vi khuẩn Gram dương lại chiếm một tỷ lệ đáng kể với 176 chủng (29,0%).
Kết quả đánh giá mức độ nhạy cảm với kháng sinh đã ghi nhận sự xuất hiện của các chủng vi khuẩn kháng kháng sinh. Đáng chú ý là Pseudomonas spp. không chỉ biểu hiện kiểu hình đa kháng mà còn cho thấy tỷ lệ kháng cao đối với cả các kháng sinh phổ rộng thế hệ ba và thế hệ bốn. Những phát hiện này cho thấy tình trạng kháng kháng sinh được phát hiện trong các chủng vi khuẩn phân lập từ mẫu cá khảo sát với mức độ khá phổ biến, điều này cho thấy sự cần thiết của việc tăng cường giám sát tình trạng kháng kháng sinh trong lĩnh vực thủy sản.
Lời cảm ơn: Các tác giả xin chân thành cảm ơn tới Viện Nghiên cứu bệnh truyền nhiễm vùng Tyumen thuộc Rospotrevnadzor, Liên bang Nga và Trung tâm Nhiệt đới Việt - Nga đã tài trợ kinh phí và hỗ trợ nhóm nghiên cứu (Đề tài M-1.6).
Tuyên bố về sử dụng Gen AI: Các tác giả khẳng định rằng không sử dụng bất kỳ công cụ AI tạo sinh nào để tạo hoặc chỉnh sửa nội dung khoa học của bản thảo này. Mọi phân tích, diễn giải và kết luận hoàn toàn do các tác giả thực hiện.
Tuyên bố về đóng góp của tác giả: Bùi Thị Thanh Nga - Phương pháp luận, viết bản thảo gốc; Posouznykh O.V., Kataeva L.V., Logacheva S.M., Stepanova T.F., Fattakhov R.G., Nguyễn Ngọc Tân - Thu thập, phân tích mẫu và xử lý số liệu; Lê Thị Lan Anh, Hoàng Đăng Hiếu - Phương pháp luận, rà soát và biên tập.
Tuyên bố về xung đột lợi ích: Tác giả bài báo này khẳng định không có bất kỳ xung đột lợi ích nào liên quan đến nghiên cứu, xuất bản bài báo này.
Tài liệu tham khảo
-
- Chen, R. Sun, C. Pan, Y. Sun, B. Mai, and Q.X. Li, Antibiotics and food safety in aquaculture, Journal of Agricultural and Food Chemistry, Vol. 68, No. 43, pp. 11908-11919, 2020. DOI:10.1021/acs.jafc.0c03996
- S. Chowdhury, et al, Antibiotics usage practices in aquaculture in Bangladesh and their associated factors, One Health, Vol. 15, 2022. DOI:10.1016/j.onehlt.2022.100445
- P. Laborda, F. Sanz-García, L. E. Ochoa-Sánchez, T. Gil-Gil, S. Hernando-Amado, and J. L. Martínez, Wildlife and antibiotic resistance, Frontiers in Cellular and Infection Microbiology, Vol. 12, 2022. DOI:10.3389/fcimb.2022.873989
- A. J. Thibodeau, M. Barret, F. Mouchet, V. X. Nguyen, and E. Pinelli, The Potential contribution of aquatic wildlife to antibiotic resistance dissemination in freshwater ecosystems: a review, Environmental Pollution, Vol. 350, 2024. DOI:10.1016/j.envpol.2024.123894
- S. E. Durand, R. Niespor, A. Ador, N. Govinda, M. Candia, and K. Torres, Ribbed mussel in an urban waterway filters bacteria introduced by sewage, Marine Pollution Bulletin, Vol. 161, 2020. DOI:10.1016/j.marpolbul.2020.111629
- A. Rico, R. Jacobs, P. J. Van den Brink, and A. Tello, A probabilistic approach to assess antibiotic resistance development risks in environmental compartments and its application to an intensive aquaculture production scenario, Environmental Pollution, Vol. 231, pp. 918–928, 2017. DOI:10.1016/j.envpol.2017.08.079
- M. L. O. Duarte, et al., In silico characterization of resistance and virulence genes in Aeromonas jandaei strains isolated from Oreochromis niloticus in Brazil, Microorganisms, Vol. 13, p. 1094, 2025. DOI: 10.3390/microorganisms13051094
- J. Zhang, et al., Mixed infection in Common carp (Cyprinus carpio) caused by Aeromonas veronii, Aeromonas hydrophila, Plesiomonas shigelloides, and Citrobacter freundii, Animals, Vol. 15, p. 805, 2025. DOI: 10.3390/ani15060805
- A. Z. Pepoyan, et al., Tetracycline resistance of Escherichia coli isolated from water, human stool, and fish gills from the Lake Sevan Basin, Letters in Applied Microbiology, Vol. 76, p. ovad021, 2023. DOI: 10.1093/lambio/ovad021
- S. T. Odonkor, K. K. Addo, Prevalence of multidrug-resistant Escherichia coli Isolated from drinking water sources, International Journal of Microbiology, 2018. DOI: 10.1155/2018/7204013
- N. A. Isokpehi, A. Beshiru, E. Green, I. H. Igbinosa, A. G. Ogofure, and E. O. Igbinosa, Characterization of Enterococcus species in surface drinking water from Akoko Edo Nigeria reveals contamination levels and risks to public health, Scientific Reports, Vol. 15, p. 38120, 2025. DOI:10.1038/s41598-025-13068-2
- A. G. Sacramento, et al., VanA-Type vancomycin-resistant Enterococcus faecium ST1336 isolated from mussels in an anthropogenically impacted ecosystem, Marine Pollution Bulletin, Vol. 142, pp. 533-536, 2019. DOI: 10.1016/j.marpolbul.2019.04.014
- D. Mohanty, et al., Prevalence of extended-spectrum β-Lactamases (ESBLs) producing Aeromonas Spp. isolated from Lamellidens marginalis (Lamark, 1819) of sewage-fed wetland: a phenotypic and genotypic approach, Microorganisms, Vol. 12, p. 723, 2024. DOI: 10.3390/microorganisms12040723
- K. Moniz, V. K. Walker, and V. Shah, Antibiotic resistance in mucosal bacteria from high arctic migratory salmonids, Environmental Microbiology Reports, Vol. 14, pp. 385-390, 2022. DOI: 10.1111/1758-2229.12975
- Huỳnh Hoàng Huy, Nguyễn Nguyễn Du, Ứng dụng GIS thể hiện sự phân bố, biến động thành phần loài và sản lượng khai thác cá vùng Đồng bằng sông Cửu Long giai đoạn 2017 - 2019, Tạp chí Nghề Cá Sông Cửu Long, số 15, pp. 83-93, tháng 12/2019.
- Vũ Đặng Hải Long, Nguyễn Đăng Quang, và Nguyễn Quang Huy, Nghiên cứu tình trạng vi khuẩn kháng kháng sinh và sự phổ biến gen kháng kháng sinh ở một số ao nuôi cá tra tại Cần Thơ và Đồng Tháp, Hội nghị Khoa học toàn quốc về Công nghệ sinh học, 2024.
- Methodological guidelines MUK 4.2.2884-11. Methods of microbiological control of environmental objects and food products. Moscow, Federal Center of Rospotrebnadzor, p. 24, 2011.
- M100Ed33, Performance Standards for antimicrobial susceptibility testing, 33rd ed. clinical & laboratory standards institute, Available online: https://clsi.org/standards/products/microbiology/documents/m100/(accessed on 10 July 2023).
- V. Jarlier, M. H. Nicolas, G. Fournier, and A. Philippon, Extended broad-spectrum beta-lactamases conferring transferable resistance to newer beta-lactam agents in Enterobacteriaceae: hospital prevalence and susceptibility patterns, Reviews of Infectious Diseases, Vol. 10, No. 4, pp. 867-78, Jul-Aug. 1988. DOI: 10.1093/clinids/10.4.867. PMID: 3263690
- A. P. Magiorakos, et al., Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance, Clinical Microbiology and Infection, Vol. 18, No. 3, pp. 268–281, Mar. 2012. DOI: 10.1111/j.1469-0691.2011.03570.x
- X. Xu, et al., Prevalence and genetic diversity of Aeromonas veronii isolated from aquaculture systems in the Poyang Lake Area, China, Frontiers in Microbiology, Vol. 13, p. 1042007, 2022. DOI: 10.3389/fmicb.2022.1042007
- R. Ehsan, et al., Aeromonas veronii isolated from climbing perch (Anabas Testudineus) suffering from Epizootic Ulcerative Syndrome (EUS), Aquaculture and Fisheries, Vol. 8, pp. 288–295, 2023.DOI: 10.1016/j.aaf.2021.11.005
- M. L. Grilo, et al., Aeromonas Spp. Prevalence, virulence, and antimicrobial resistance in an ex situ program for threatened freshwater fish: a pilot study with protective measures, Animals, Vol. 12, p. 436, 2022. DOI: 10.3390/ani12040436
- A. Lupo, M. Haenni, and J. Y. Madec, Antimicrobial resistance in Acinetobacter spp. and Pseudomonas spp, Microbiology Spectrum, Vol. 6, 2017. DOI: 10.1128/microbiolspec.arba-0007-2017
- K. M. G. M. M. Kariyawasam, N. K. Lee, and H. D. Paik, Fermented dairy products as delivery vehicles of novel probiotic strains isolated from traditional fermented asian foods, Journal of Food Science and Technology, Vol. 58, pp. 2467–2478, 2021. DOI: 10.1007/s13197-020-04857-w
- A. A. Adegoke, C. E. Madu, P. Reddy, O. K. Fatunla,T. A. Stenström, and A. I. Okoh, Virulence and antibiotic resistance genes in Enterococcus from wastewater for reuse and their health impact, Microorganisms, Vol. 13, p. 1045, 2025. DOI: 10.3390/microorganisms13051045
- A. Sampaio, V. Silva, P. Poeta, and F. Aonofriesei, Vibrio Spp.: life strategies, ecology, and risks in a changing environment, Diversity, Vol. 14, p. 97, 2022. DOI: 10.3390/d14020097
- F. Di Bartolomeo, R. Ligresti, S. Pettenuzzo, T. Bini, C. Tincati, and G. C. Marchetti, Shewanella putrefaciens, an emerging foe from climate change: a case report, Journal of Medical Case Reports, vol. 19, p. 105, 2025. DOI:10.1186/s13256-025-05100-w
- Z. Pang, R. Raudonis, B. R. Glick, T. J. Lin, and Z. Cheng, Antibiotic resistance in Pseudomonas aeruginosa: mechanisms and alternative therapeutic strategies, Biotechnology Advances, Vol. 37, pp. 177–192, 2019. DOI:10.1016/j.biotechadv.2018.11.013
- G. Tebano, I. Zaghi, M. Cricca, and F. Cristini, Antibiotic treatment of infections caused by AmpC-producing Enterobacterales, Pharmacy, Vol. 12, p. 142, 2024. DOI:10.3390/pharmacy12050142
- I. Ferheen, R. Spurio, L. Mancini, and S. Marcheggiani, Detection of morganella morganii bound to a plastic substrate in surface water, Journal of Global Antimicrobial Resistance, Vol. 32, pp. 104–107, 2023. DOI:10.1016/j.jgar.2023.01.008
- N. T. Nhung, N. V. Cuong, G. Thwaites, and J. Carrique-Mas, Antimicrobial usage and antimicrobial resistance in animal production in Southeast Asia: a review, Antibiotics, Vol. 5, p. 37, 2016. DOI:10.3390/antibiotics5040037
- K. Bush, P. A. Bradford, β-Lactams and β-Lactamase inhibitors: an overview, Cold Spring Harbor Perspectives in Medicine, Vol. 6, p. a025247, 2016. DOI:10.1101/cshperspect.a025247
- F. C. Uhland., et al., Extended spectrum β-Lactamase-producing enterobacterales of shrimp and salmon available for purchase by consumers in Canada-a risk profile using the codex framework, Antibiotics, Vol. 12, p. 1412, 2023. DOI:10.3390/antibiotics12091412