Đánh giá tác động của Cu²⁺ lên đáp ứng stress oxy hóa và sửa chữa DNA thông qua biểu hiện gene và protein trong phôi Cá ngựa vằn (Danio rerio)
Trường Đại học Lạc Hồng, Đồng Nai, Việt Nam
Đồng Nai, Việt Nam
Email: khoadd@lhu.edu.vn
Nội dung chính của bài viết
Tóm tắt
Ô nhiễm kim loại nặng, đặc biệt là Cu²⁺, trong môi trường nước là mối quan tâm toàn cầu. Nghiên cứu này nhằm mục đích đánh giá một cách có hệ thống các đáp ứng phân tử của phôi cá ngựa vằn (Danio rerio) khi tiếp xúc với các nồng độ Cu²⁺ khác nhau (0,1; 1,0 và 10,0 µg/L Cu²⁺) trong 24 giờ. Nghiên cứu tập trung vào các dấu ấn sinh học chính liên quan đến giải độc kim loại, đáp ứng tổn thương DNA và chống oxy hóa, bao gồm metallothionein 2 (mt2), các protein GADD45 (GADD45A, GADD45G) và các enzyme chống oxy hóa superoxide dismutase (SOD1, SOD2). Phương pháp Real-time PCR (qPCR) và Western blot được sử dụng đồng thời để phân tích cả mức độ phiên mã và dịch mã. Kết quả cho thấy mức biểu hiện của các gene (mt2, gadd45a, gadd45g, sod1, sod2) và các protein tương ứng (MT2, GADD45A, GADD45G, SOD1, SOD2) có đáp ứng rõ ràng theo nồng độ Cu²⁺. Ở nồng độ 10 µg/L Cu²⁺, biểu hiện của các gene mt2, gadd45g, sod1 và sod2 tăng mạnh ở mức độ mRNA (gấp 9,1 đến 17,5 lần), đồng thời mức protein tương ứng (MT2, GADD45G, SOD1 và SOD2) cũng tăng đáng kể (gấp 8,5 đến 15,8 lần). Ngược lại, gadd45a bị ức chế rõ rệt (giảm 11 lần ở mức độ mRNA), trong khi protein tương ứng (GADD45A) cũng giảm đáng kể (6,7 lần) ở nhóm 1,0 µg/L Cu²⁺. Sự tương đồng trong xu hướng biểu hiện gene và protein cho thấy các cơ chế đáp ứng chủ yếu được điều hòa ở mức độ phiên mã. Sự điều hòa trái ngược giữa GADD45A và GADD45G phản ánh một chiến lược thích nghi của tế bào nhằm cân bằng giữa sửa chữa DNA, sống sót và chết tế bào. Nghiên cứu này cung cấp dữ liệu nền tảng quan trọng cho đánh giá rủi ro sinh thái và sự hiểu biết sâu sắc hơn về cơ chế độc tính của Cu²⁺ ở cấp độ phân tử, đồng thời khẳng định vai trò của cá ngựa vằn như một mô hình sinh học nhạy cảm để giám sát ô nhiễm.
Abstract
Effects of Cu²⁺ Exposure on Oxidative Stress and DNA Damage Response via Gene and Protein Expression in Zebrafish (Danio rerio) Embryos
Heavy metal pollution, particularly copper (Cu²⁺), in aquatic environments is a global concern. This study aimed to systematically evaluate the molecular responses of zebrafish (Danio rerio) embryos exposed to different Cu²⁺ concentrations (0.1, 1.0, and 10.0 µg/L Cu²⁺) for 24 hours. We focused on key genes and proteins involved in metal detoxification (MT2), DNA damage response (GADD45A, GADD45G), and antioxidant defense (SOD1, SOD2). Real-time PCR (qPCR) and Western blot were used in parallel to analyze both transcriptional and translational levels. The results revealed a clear concentration-dependent response. At 10 µg/L Cu²⁺, the expression of MT2, GADD45G, SOD1, and SOD2 increased significantly at both mRNA (9.1–17.5-fold) and protein (8.5–15.8-fold) levels. Conversely, GADD45A showed significant inhibition (11-fold decrease in mRNA and 6.7-fold decrease in protein at the 1.0 µg/L Cu²⁺ exposure level). The consistent trends observed between gene and protein expression suggest that the primary response mechanisms are largely regulated at the transcriptional level. The contrasting regulation of GADD45A and GADD45G indicates a sophisticated cellular strategy to balance DNA repair, survival, and cell death. These findings provide important insights into the molecular mechanisms of copper toxicity and highlight the potential of MT2, SOD2, and GADD45G as sensitive molecular biomarkers for monitoring Cu²⁺ contamination in aquatic environments.
Keywords: Biomarker; copper toxicity; DNA damage; gene expression; oxidative stress; Western blot;zebrafish
Từ khóa
Độc tính Cu²⁺, cá ngựa vằn, stress oxy hóa, tổn thương DNA, biểu hiện gene, Western blot, dấu ấn sinh học
Chi tiết bài viết

Bài báo này được cấp phép theo Creative Commons Attribution 4.0 International License.
Điểm nổi bật
Phát hiện chính: Cu²⁺ gây ra sự điều hòa trái ngược giữa hai gene sửa chữa DNA: gadd45a bị ức chế (giảm 11 lần) trong khi gadd45g được cảm ứng mạnh (tăng 17.5 lần).
Tác động rõ rệt: Ở nồng độ cao (10 µg/L Cu²⁺), các gene giải độc mt2 và chống oxy hóa (sod1, sod2) tăng mạnh theo nồng độ (gấp 9-17 lần).
Giá trị thực tiễn: Bộ ba MT2, SOD2 và GADD45G là các dấu ấn sinh học tiềm năng để giám sát ô nhiễm Cu²⁺ trong môi trường nước
1. ĐẶT VẤN ĐỀ
Sự phát triển nhanh chóng của đô thị hóa và công nghiệp hóa đã làm gia tăng sự tích tụ kim loại nặng trong môi trường nước, gây rủi ro đối với hệ sinh thái và sức khỏe con người [1, 2]. Các nguồn phát thải chính bao gồm nước thải công nghiệp (luyện kim, điện tử), khí thải từ nhà máy nhiệt điện và phương tiện giao thông, cũng như các hoạt động khai thác mỏ [2, 3]. Trong số các kim loại nặng, đồng (Cu²⁺) là một nguyên tố vi lượng thiết yếu, đóng vai trò là đồng yếu tố thiết yếu trong nhiều quá trình sinh lý như hô hấp tế bào và hệ thống chống oxy hóa [3].
Tuy nhiên, khi vượt quá ngưỡng sinh học, Cu²⁺ trở nên độc hại do khả năng xúc tác hình thành các loại oxy phản ứng (ROS) thông qua các phản ứng oxy hóa khử, dẫn đến stress oxy hóa và tổn thương protein, lipid cũng như DNA.
Cá ngựa vằn (Danio rerio) là một mô hình lý tưởng trong nghiên cứu độc chất học môi trường và sinh học phát triển nhờ các đặc điểm như kích thước nhỏ, sinh sản nhanh, phôi phát triển ngoài cơ thể và trong suốt, cùng với mức độ tương đồng di truyền cao với người [4, 5]. Ngoài ra, các con đường đáp ứng stress ở cá ngựa vằn được bảo tồn cao, giúp loài này trở thành một hệ thống cảnh báo sinh học hiệu quả đối với ô nhiễm môi trường nước [5, 6]. Để đối phó với stress kim loại, tế bào phát triển một mạng lưới phòng vệ phức tạp. Trong đó, metallothionein 2 (MT2) là một protein liên kết kim loại có vai trò quan trọng trong giải độc kim loại và điều hòa stress oxy hóa [6].
Các protein thuộc họ GADD45 (GADD45A và GADD45G) đóng vai trò quan trọng trong đáp ứng tổn thương DNA, tham gia điều hòa chu kỳ tế bào, sửa chữa DNA và apoptosis [7–9]. Sự điều hòa khác biệt giữa các thành viên trong họ này có thể quyết định số phận tế bào, từ sửa chữa đến chết theo chương trình [7]. Quá trình apoptosis - một cơ chế sinh học cơ bản trong đáp ứng stress tế bào - đóng vai trò quan trọng trong duy trì cân bằng nội môi [10]. Hệ enzyme superoxide dismutase (SOD1 và SOD2) đóng vai trò tuyến phòng thủ đầu tiên chống lại stress oxy hóa, xúc tác chuyển đổi superoxide thành hydrogen peroxide và oxy [11,12].
Mặc dù đã có nhiều nghiên cứu về độc tính của kim loại nặng, các nghiên cứu đánh giá đồng thời mối tương quan giữa biểu hiện gene và protein của các dấu ấn sinh học này trong phôi cá ngựa vằn ở giai đoạn phát triển sớm vẫn còn rất hạn chế. Do đó, nghiên cứu này nhằm: (1) đánh giá tác động của Cu²⁺ lên biểu hiện của các gene mt2, gadd45a, gadd45g, sod1 và sod2; (2) phân tích mối tương quan giữa các đáp ứng ở mức độ phiên mã và dịch mã; và (3) cung cấp cơ sở khoa học cho việc ứng dụng các dấu ấn sinh học này trong giám sát ô nhiễm Cu²⁺ trong môi trường nước [13].
2. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Nuôi cá và thu phôi
Cá ngựa vằn trưởng thành (dòng AB) được duy trì trong hệ thống tuần hoàn dưới các điều kiện tiêu chuẩn: nhiệt độ 28 ± 1°C, pH 7,2 ± 0,2, độ dẫn điện 500 µS/cm và chu kỳ sáng/tối 14:10 giờ. Cá được cho ăn hai lần mỗi ngày bằng artemia và thức ăn viên công nghiệp. Để thu phôi, cá bố mẹ được cho giao phối tự nhiên vào sáng sớm với tỷ lệ đực:cái là 1:2. Phôi được thu thập trong vòng 30 phút sau khi đẻ, rửa sạch và chọn lọc những phôi phát triển bình thường (giai đoạn phôi nang sớm) dưới kính hiển vi soi nổi (stereomicroscope)
2.2. Bố trí thí nghiệm
Phôi cá ngựa vằn được phân bố ngẫu nhiên hoàn toàn vào bốn nhóm xử lý (150 phôi mỗi nhóm cho mỗi lần lặp lại (n = 3): (1) nhóm đối chứng được nuôi trong môi trường tiêu chuẩn (E3); (2) nhóm xử lý với 0,1 µg/L Cu²⁺; (3) nhóm xử lý với 1,0 µg/L Cu²⁺; và (4) nhóm xử lý với 10,0 µg/L Cu²⁺.
Các dung dịch xử lý được chuẩn bị từ CuSO₄·5H₂O (độ tinh khiết ≥ 99%, cấp phân tích). Nồng độ đồng trong thí nghiệm được biểu diễn theo hàm lượng ion Cu²⁺ tương đương (µg/L Cu²⁺). Các giá trị này được tính toán dựa trên khối lượng phân tử của CuSO₄·5H₂O để đảm bảo tính chính xác và khả năng so sánh với các nghiên cứu trước.
Phôi được nuôi trong đĩa Petri polystyrene đường kính 90 mm chứa 50 mL môi trường tương ứng và ủ ở 28 ± 1°C trong 24 giờ. Môi trường được thay mới sau 12 giờ nhằm duy trì nồng độ Cu²⁺ ổn định trong suốt thời gian thí nghiệm. pH của môi trường được theo dõi và duy trì ổn định trong suốt quá trình thí nghiệm.
2.3. Phân tích biểu hiện gene bằng real-time PCR (qPCR)
Tổng RNA được tách chiết từ phôi cá ngựa vằn bằng bộ kit TRIzol Reagent (Thermo Scientific, USA) theo hướng dẫn của nhà sản xuất. Nồng độ và độ tinh sạch của RNA được xác định bằng máy NanoDrop 2000 spectrophotometer (Thermo Scientific, USA), với tỷ lệ A260/A280 trong khoảng 1,8–2,0.
Thư viện cDNA được tổng hợp từ 1 µg RNA tổng số bằng bộ kit RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific, USA) theo quy trình của nhà sản xuất.
Phản ứng qPCR được thực hiện trên hệ thống QuantStudio 5 (Applied Biosystems, USA) sử dụng bộ kit SYBR Green qPCR Master Mix (PCR Biosystems, UK). Quy trình qPCR được thực hiện theo phương pháp 2 bước (two-step), sử dụng cDNA làm khuôn. Thể tích phản ứng là 20 µL, bao gồm 10 µL master mix, 0,8 µL mồi xuôi và mồi ngược (10 µM), 2 µL cDNA và 6,4 µL nước không chứa nuclease. Trình tự mồi sử dụng trong real-time PCR (Bảng 1)
Chu trình nhiệt gồm: 95°C trong 2 phút, tiếp theo là 40 chu kỳ gồm 95°C trong 10 giây, 58°C trong 15 giây và 72°C trong 15 giây. Độ đặc hiệu của sản phẩm PCR được kiểm tra bằng phân tích đường cong nóng chảy (melting curve analysis). Biểu hiện gene được chuẩn hóa theo gene nội chuẩn ef1α và tính toán theo phương pháp 2^-ΔΔCt [14].
Bảng 1. Trình tự mồi sử dụng trong real-time PCR (qPCR)
Gene | Trình tự mồi xuôi (forward) (5' - 3') | Trình tự mồi ngược (reverse) (5' - 3') | Kích thước sản phẩm (bp) |
mt2 | CCT GCA AGT GCA CTA ATT GCC AGT | TCT GTT TCA AGA AGC CGA AAG CCC | 120 |
gadd45a | AAC GTG GTC TTG TGT CTG CT | AGG TCC ATC GAC TCT CCT CC | 152 |
gadd45g | CGC CTT GGA TAC GTC CG | CTC TTG ACA CGC GAC CAG TA | 145 |
sod1 | TGA GAC ACG TCG GAG ACC | TGC CGA TCA CTC CAC AGG | 138 |
sod2 | TTC AGG GCT CAG GCT GG | ATG GCT TTA ACA TAG TCC GGT | 161 |
ef1α
| GTA CTA CTC TTC TTG ATG CCC | GTA CAG TTC CAA TAC CCT CCA | 110 |
Nguồn: Mồi cho mt2, sod1, sod2 được điều chỉnh từ [15]; mồi cho gadd45a, gadd45g được điều chỉnh từ [14]; mồi ef1α từ [14, 15].
2.4. Tách chiết protein và phân tích Western blot
Một nhóm phôi song song (50 phôi cho mỗi mẫu, n = 3) được sử dụng để phân tích protein. Mẫu được đồng nhất hóa trong 200 µL đệm ly giải RIPA (Beyotime, China) có chứa 1 mM PMSF và hỗn hợp ức chế protease. Dịch đồng nhất được ly tâm ở 12.000 × g trong 15 phút ở 4°C. Protein hòa tan trong phần nổi phía trên được thu thập và định lượng bằng phương pháp Bradford sử dụng albumin huyết thanh bò (BSA) làm chuẩn [16]. 30 µg protein từ mỗi mẫu được điện di trên gel SDS-PAGE 12% và sau đó chuyển lên màng PVDF (Millipore, USA). Màng được ủ trong dung dịch chặn (5% sữa gầy Tris-buffered saline containing 0,1% Tween-20 (TBST)) trong 2 giờ ở nhiệt độ phòng. Tiếp theo, màng được ủ qua đêm ở 4°C với các kháng thể sơ cấp sau: anti-MT2 (Abcam, ab12228, pha loãng 1:1000), anti-GADD45A (Cell Signaling Technology, #4632, 1:1000), anti-GADD45G (Santa Cruz, sc-133685, 1:500), anti-SOD1 (Abcam, ab13498, 1:2000), anti-SOD2 (CST, #13141, 1:1000), và anti-β-Actin (Sigma, A5441, 1:5000) làm đối chứng nội sinh. Sau khi rửa, màng được ủ với kháng thể thứ cấp liên hợp HRP (goat anti-rabbit IgG hoặc goat anti-mouse IgG, pha loãng 1:5000) trong 2 giờ ở nhiệt độ phòng (~25°C). Tín hiệu được phát hiện bằng thuốc thử ECL Prime (Cytiva, USA) và chụp ảnh trên hệ thống ChemiDoc™ MP Imaging System (Bio-Rad, USA). Cường độ vạch được phân tích định lượng bằng phần mềm Image Lab (Bio-Rad, USA) và chuẩn hóa theo β-actin.
2.5. Phân tích thống kê
Các thí nghiệm được thực hiện với ba lần lặp lại sinh học độc lập (biological replicates). Dữ liệu được trình bày dưới dạng giá trị trung bình ± độ lệch chuẩn (Mean ± SD). Sự khác biệt giữa các nhóm được phân tích bằng phương pháp phân tích phương sai một yếu tố (One-way ANOVA) và kiểm định hậu kiểm Tukey cho so sánh nhiều cặp. Giá trị p < 0.05 được coi là có ý nghĩa thống kê. Tất cả các phân tích được thực hiện bằng phần mềm GraphPad Prism phiên bản 9.0 (GraphPad Software, USA).
3. KẾT QUẢ
3.1. Tác động của Cu²⁺ lên biểu hiện gene
Kết quả phân tích qPCR cho thấy Cu²⁺ gây ra những thay đổi mạnh mẽ và phụ thuộc vào nồng độ trong biểu hiện của tất cả các gene được nghiên cứu sau 24 giờ xử lý (Bảng 2, Hình 1).
Bảng 2. Mức độ biểu hiện tương đối của các gene mt2, gadd45a, gadd45g, sod1 và sod2 trong phôi cá ngựa vằn 24 hpf được xử lý với các nồng độ Cu²⁺ khác nhau (0,1; 1,0; 10,0 µg/L Cu²⁺)
Gene | Đối chứng | 0,1 µg/L | 1,0 µg/L | 10,0 µg/L |
mt2 | 1,00 ± 0,10 | 1,15 ± 0,12 | 1,20 ± 0,15 | 9,10 ± 0,95* |
gadd45a | 1,00 ± 0,08 | 0,63 ± 0,06* | 0,09 ± 0,01* | 0,15 ± 0,02* |
gadd45g | 1,00 ± 0,09 | 1,50 ± 0,14 | 2,30 ± 0,20* | 17,50 ± 1,80* |
sod1 | 1,00 ± 0,11 | 1,25 ± 0,13 | 1,40 ± 0,16 | 14,20 ± 1,50* |
sod2 | 1,00 ± 0,10 | 1,30 ± 0,12 | 1,50 ± 0,14 | 14,50 ± 1,55* |
*p < 0,05 so sánh cụ thể với nhóm đối chứng
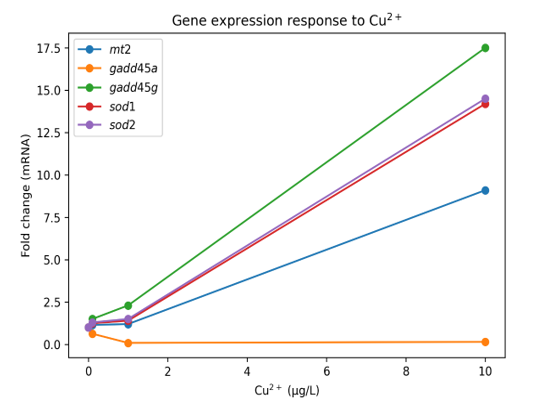
Hình 1. Biểu hiện tương đối của các gene mt2, gadd45a, gadd45g, sod1 và sod2 theo nồng độ Cu²⁺.
Biểu hiện gene mt2 tăng mạnh gấp 9,1 lần ở nồng độ 10 µg/L Cu²⁺ so với đối chứng (p < 0,05). Ở các nồng độ thấp hơn (0,1 và 1,0 µg/L Cu²⁺ ), sự thay đổi không có ý nghĩa.
Gene gadd45a: Ngược lại, gene gadd45a bị ức chế sâu sắc. Ở nồng độ 1,0 µg/L Cu²⁺ , biểu hiện gene giảm mạnh nhất, chỉ còn 9% so với mức đối chứng (giảm 11 lần, p < 0,05). Ở các nồng độ khác, biểu hiện gene cũng giảm đáng kể.
Gene gadd45g cho thấy xu hướng biểu hiện hoàn toàn trái ngược với gene gadd45a. Biểu hiện gene tăng dần theo nồng độ, đạt mức tăng gấp 17,5 lần ở nhóm 10 µg/L Cu²⁺ (p < 0,05).
Các gene sod1 và sod2 đều cho thấy xu hướng biểu hiện tăng mạnh, có ý nghĩa chỉ ở nồng độ cao nhất (10 µg/L Cu²⁺), với mức tăng lần lượt là 14,2 và 14,5 lần (p < 0,05). Ở các nồng độ thấp hơn, sự thay đổi không có ý nghĩa thống kê.
3.2. Tác động của đồng Cu²⁺ lên biểu hiện protein
Kết quả phân tích định lượng protein bằng phương pháp Western blot (Bảng 3, Hình 2) cho thấy xu hướng biểu hiện protein nhìn chung tương tự với dữ liệu phiên mã, mặc dù có sự khác biệt về mức độ.
Protein MT2 tăng mạnh ở nồng độ 10 µg/L Cu²⁺ (gấp 8,5 lần so với đối chứng, p < 0,05), trong khi ở các nồng độ thấp hơn sự thay đổi không có ý nghĩa thống kê.
Protein GADD45A giảm rõ rệt, đặc biệt ở nhóm 1,0 µg/L Cu²⁺ (giảm 6,7 lần), phù hợp với xu hướng giảm ở mức độ mRNA
Bảng 3. Mức độ biểu hiện protein tương đối của MT2, GADD45A, GADD45G, SOD1 và SOD2 trong phôi cá ngựa vằn 24 hpf được xử lý với các nồng độ Cu²⁺
Protein | Đối chứng | 0,1 µg/L Cu²⁺ | 1,0 µg/L Cu²⁺ | 10,0 µg/L Cu²⁺ |
MT2 | 1,00 ± 0,12 | 1,30 ± 0,15 | 1,45 ± 0,18 | 8,50 ± 0,90* |
GADD45A | 1,00 ± 0,10 | 0,70 ± 0,08 | 0,15 ± 0,02* | 0,25 ± 0,03* |
GADD45G | 1,00 ± 0,11 | 1,40 ± 0,16 | 2,10 ± 0,25* | 15,80 ± 1,65* |
SOD1 | 1,00 ± 0,09 | 1,35 ± 0,14 | 1,60 ± 0,17* | 12,50 ± 1,30* |
SOD2 | 1,00 ± 0,13 | 1,25 ± 0,15 | 1,55 ± 0,18 | 13,50 ± 1,40* |
*p < 0,05 so sánh cụ thể với nhóm đối chứng
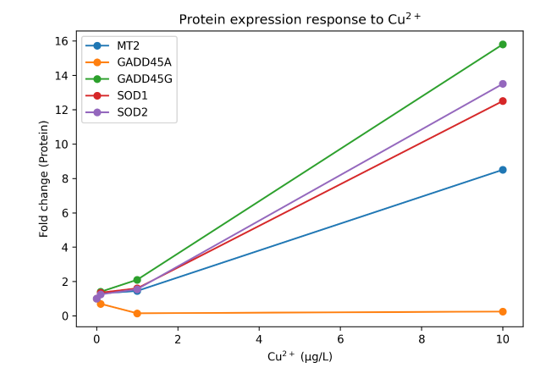
Hình 2. Biểu hiện protein MT2, GADD45A, GADD45G, SOD1 và SOD2 theo nồng độ Cu²⁺.
Ngược lại, protein GADD45G tăng mạnh theo nồng độ và đạt mức cao nhất ở nhóm 10 µg/L Cu²⁺ (tăng 15,8 lần, p < 0,05).
Protein SOD1 và SOD2 đều tăng đáng kể ở nồng độ cao nhất, với mức tăng lần lượt là 12,5 và 13,5 lần (p < 0,05).
4. THẢO LUẬN
Nghiên cứu này cung cấp một cái nhìn toàn diện về đáp ứng phân tử của phôi cá ngựa vằn đối với stress do Cu²⁺ gây ra thông qua phân tích tích hợp giữa mức độ phiên mã và dịch mã. Kết quả cho thấy cơ chế đáp ứng phức tạp, phụ thuộc nồng độ và đặc hiệu theo từng gene/protein.
4.1. Đáp ứng giải độc và chống oxy hóa
Sự gia tăng mạnh mẽ biểu hiện của protein MT2 ở nồng độ 10 µg/L Cu²⁺ là một đáp ứng điển hình đối với stress kim loại nặng và phù hợp với các nghiên cứu trước đây [6]. Sự cảm ứng này có liên quan đến hoạt hóa các cơ chế điều hòa nội bào nhằm duy trì cân bằng kim loại. MT2 có khả năng liên kết với các ion Cu²⁺ tự do, góp phần làm giảm độc tính và hạn chế sự tham gia của Cu²⁺ vào các phản ứng oxy hóa khử tạo ROS.
Sự gia tăng đồng thời của protein SOD1 và SOD2 cho thấy sự hiện diện rõ rệt của stress oxy hóa. Đặc biệt, SOD2 đóng vai trò quan trọng trong ty thể - nơi sản sinh chính các gốc superoxide trong quá trình phosphoryl hóa oxy hóa. Kết quả này phù hợp với các nghiên cứu trước đó về stress oxy hóa và độc tính kim loại trên cá ngựa vằn [11,12, và 17]. Ngoài ra, các nghiên cứu gần đây cũng cho thấy stress oxy hóa và tổn thương ty thể là cơ chế trung tâm trong độc tính kim loại kết hợp và các tác nhân môi trường khác [18].
4.2. Đáp ứng tổn thương DNA và sự điều hòa trái ngược của GADD45
Một phát hiện quan trọng của nghiên cứu là sự điều hòa trái ngược giữa hai protein GADD45A và GADD45G. Dữ liệu cho thấy GADD45A giảm đáng kể, đặc biệt ở nồng độ 1,0 µg/L Cu²⁺, trong khi GADD45G lại tăng mạnh theo nồng độ.
Theo Hollander và cộng sự (1993) [8], GADD45A là một thành phần quan trọng trong đáp ứng tổn thương DNA và có liên quan đến quá trình apoptosis. Sự giảm biểu hiện của GADD45A trong nghiên cứu này có thể phản ánh một cơ chế thích nghi nhằm hạn chế kích hoạt sớm quá trình chết tế bào theo chương trình, một hiện tượng sinh học cơ bản đã được mô tả bởi Kerr và cộng sự (1972) [10].
Ngược lại, GADD45G được cảm ứng mạnh, phù hợp với vai trò điều hòa chu kỳ tế bào và hỗ trợ sửa chữa DNA đã được báo cáo trước đó [9]. Sự gia tăng này có thể cho phép tế bào kéo dài thời gian sửa chữa tổn thương trước khi tiếp tục chu kỳ tế bào. Ngoài ra, các protein thuộc họ GADD45 đã được chứng minh có vai trò như các cảm biến stress phân tử trong các cơ chế sửa chữa DNA [7].
Sự điều hòa trái ngược giữa GADD45A và GADD45G cho thấy một chiến lược thích nghi tinh vi của tế bào: ở mức stress trung bình, tế bào ưu tiên hạn chế apoptosis để duy trì sự sống sót; trong khi ở mức stress cao hơn, các cơ chế sửa chữa DNA được tăng cường nhằm bảo vệ tính toàn vẹn bộ gene.
4.3. Ý nghĩa sinh thái và môi trường
Kết quả nghiên cứu không chỉ cung cấp hiểu biết về cơ chế mà còn có giá trị ứng dụng trong giám sát môi trường. Các protein MT2, SOD2 và GADD45G cho thấy đáp ứng rõ ràng và phụ thuộc nồng độ đối với Cu²⁺, cho thấy tiềm năng sử dụng như các dấu ấn sinh học phân tử.
Việc kết hợp nhiều dấu ấn sinh học (ví dụ: MT2 phản ánh phơi nhiễm kim loại, SOD2 phản ánh stress oxy hóa, và GADD45G phản ánh tổn thương DNA) có thể cung cấp một đánh giá toàn diện hơn về mức độ ô nhiễm so với chỉ dựa vào nồng độ Cu²⁺ trong môi trường nước.
5. KẾT LUẬN
Nghiên cứu này làm sáng tỏ tác động của Cu²⁺ lên đáp ứng phân tử của phôi cá ngựa vằn thông qua phân tích tích hợp giữa biểu hiện gene và protein. Kết quả cho thấy Cu²⁺ gây ra các thay đổi phụ thuộc nồng độ trong biểu hiện của các gene mt2, gadd45a, gadd45g, sod1 và sod2, đồng thời phản ánh tương ứng ở mức độ protein.
Sự gia tăng mạnh của các protein MT2, SOD1 và SOD2 xác nhận vai trò quan trọng của các cơ chế giải độc kim loại và chống oxy hóa trong đáp ứng với stress do Cu²⁺. Ngược lại, sự điều hòa trái ngược giữa GADD45A và GADD45G cho thấy một chiến lược thích nghi phức tạp của tế bào nhằm cân bằng giữa sửa chữa DNA và kiểm soát chết tế bào.
Các protein MT2, SOD2 và GADD45G cho thấy đáp ứng rõ ràng và phụ thuộc nồng độ đối với Cu²⁺, qua đó thể hiện tiềm năng sử dụng như các dấu ấn sinh học phân tử trong giám sát ô nhiễm Cu²⁺ trong môi trường nước.
Đạo đức nghiên cứu: Nghiên cứu sử dụng phôi Cá ngựa vằn (Danio rerio) được thực hiện theo các quy định hiện hành về sử dụng động vật trong nghiên cứu khoa học và đảm bảo giảm thiểu tối đa đau đớn cho sinh vật thí nghiệm.
Lời cảm ơn và nguồn tài trợ: Nghiên cứu này không nhận tài trợ từ bất kỳ tổ chức nào.
Tuyên bố về việc sử dụng trí tuệ nhân tạo (AI): Nhóm tác giả có sử dụng công cụ trí tuệ nhân tạo (AI) chỉ nhằm hỗ trợ chỉnh sửa ngôn ngữ và trình bày bản thảo. Nội dung khoa học, phân tích dữ liệu và kết luận hoàn toàn do các tác giả thực hiện và chịu trách nhiệm.
Tuyên bố về đóng góp của tác giả: Ngô Văn Tuấn:Thiết kế nghiên cứu, xây dựng ý tưởng, phân tích dữ liệu và viết bản thảo và chỉnh sửa bản thảo; Đặng Đăng Khoa: Thực hiện thí nghiệm, thu thập dữ liệu, hỗ trợ phân tích và chỉnh sửa bản thảo; Nguyễn Thi Thu Hằng: Xử lý số liệu, hỗ trợ viết và chỉnh sửa bản thảo. Tất cả các tác giả đã đọc, góp ý và đồng ý với phiên bản cuối cùng của bài báo.
Tuyên bố về xung đột lợi ích: Các tác giả cam kết không có xung đột lợi ích liên quan đến nghiên cứu này.
Tài liệu tham khảo
- J. O. Duruibe, M. O. C. Ogwuegbu và J. N. Egwurugwu, Heavy metal pollution and human biotoxic effects, International Journal of Physical Sciences, Vol. 2, No. 5, pp. 112–118, 2007.
- G. Ziemacki, G. Viviano và F. Merli, Heavy metals: sources and environmental presence, Annali dell’Istituto Superiore di Sanità, Vol. 25, No. 3, pp. 531–536, 1989.
- M. C. Linder, M. Hazegh-Azam, Copper biochemistry and molecular biology, American Journal of Clinical Nutrition, Vol. 63, No. 5, pp. 797S–811S, 1996. DOI: 10.1093/ajcn/63.5.797
- C. B. Kimmel, W. W. Ballard, S. R. Kimmel, B. Ullmann và T. F. Schilling, Stages of embryonic development of the zebrafish, Developmental Dynamics, Vol. 203, No. 3, pp. 253–310, 1995. DOI: 10.1002/aja.1002030302
- S. Scholz, S. Fischer, U. Gündel, E. Küster, T. Luckenbach và D. Völker, The zebrafish embryo model in environmental risk assessment—applications beyond acute toxicity testing, Environmental Science and Pollution Research, Vol. 15, No. 5, pp. 394–404, 2008. DOI: 10.1007/s11356-008-0018-z
- J. M. Rice, A. Zweifach và M. A. Lynes, Metallothionein regulates intracellular zinc signaling during CD4+ T cell activation, BMC Immunology, Vol. 17, Art. 13, 2016. DOI: 10.1186/s12865-016-0151-2
- A. Schäfer, Gadd45 proteins: key players of repair-mediated DNA demethylation, Advances in Experimental Medicine and Biology, Vol. 793, pp. 35–50, 2013. DOI: 10.1007/978-1-4614-8289-5_3
- M. C. Hollander, I. Alamo, J. Jackman, M. G. Wang, O. W. McBride và A. J. Fornace Jr., Analysis of the mammalian Gadd45 gene and its response to DNA damage, Journal of Biological Chemistry, Vol. 268, No. 32, pp. 24385–24393, 1993.
- K. Ishida, Y. Yuge, M. Hanaoka, N. Iwamoto, S. Masuda và O. Ohazama, Gadd45g regulates dental epithelial cell proliferation through p38 MAPK-mediated p21 expression, Genes to Cells, Vol. 18, No. 8, pp. 660–671, 2013. DOI: 10.1111/gtc.12067
- J. F. R. Kerr, A. H. Wyllie và A. R. Currie, Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics, British Journal of Cancer, Vol. 26, No. 4, pp. 239–257, 1972. DOI: 10.1038/bjc.1972.33
- X. Liu et al., Sirt3-dependent deacetylation of SOD2 plays a protective role against oxidative stress in oocytes from diabetic mice, Cell Cycle, Vol. 16, No. 13, pp. 1302–1308, 2017. DOI: 10.1080/15384101.2017.1320004
- K. Park et al., Effects of thermal stress-induced lead (Pb) toxicity on apoptotic cell death, inflammatory response, oxidative defense, and DNA methylation in zebrafish (Danio rerio) embryos, Aquatic Toxicology, Vol. 224, Art. 105479, 2020. DOI: 10.1016/j.aquatox.2020.105479
- W. Huang et al., A transcriptomics-based analysis of toxicity mechanisms of zebrafish embryos and larvae following parental bisphenol A exposure, Ecotoxicology and Environmental Safety, Vol. 205, Art. 111165, 2020. DOI: 10.1016/j.ecoenv.2020.111165
- K. J. Livak, T. D. Schmittgen, Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method, Methods, Vol. 25, No. 4, pp. 402–408, 2001. DOI: 10.1006/meth.2001.1262
- S. Sakthivel et al., Molecular characterization and expression profiling of arsenic mediated stress-responsive genes in Dawkinsia tambraparniei, Journal of Applied Biology & Biotechnology, Vol. 11, No. 7, pp. 193–199, 2023. DOI: 10.7324/JABB.2023.117527
- M. M. Bradford, A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding, Analytical Biochemistry, Vol. 72, No. 1–2, pp. 248–254, 1976. DOI: 10.1016/0003-2697(76)90527-3
- V. Curcio et al., Morphological and molecular alterations induced by lead in embryos and larvae of Danio rerio, Applied Sciences, Vol. 11, No. 16, Art. 7464, 2021. DOI: 10.3390/app11167464
- K. Dang et al., Combined effects of microplastics and cadmium on oxidative stress, DNA damage, and embryonic development in zebrafish (Danio rerio), FishTaxa, Vol. 37, pp. 78–86, 2025.