Tăng trưởng và tỷ lệ sống của hai loài san hô thuộc giống Acropora (A. muricata và A. robusta) trên các giá thể khác nhau tại Trạm thử nghiệm Đầm Báy, Vịnh Nha Trang
Chi nhánh Ven Biển, Trung tâm Nhiệt đới Việt - Nga, Khánh Hòa, Việt Nam
Số 30 Nguyễn Thiện Thuật, Nha Trang, Khánh Hòa, Việt Nam
Email: mrtran_cnvb@yahoo.com
Nội dung chính của bài viết
Tóm tắt
Sự suy giảm nghiêm trọng quần thể san hô, đặc biệt ở các loài thuộc giống Acropora, đã nhấn mạnh nhu cầu cấp thiết của công tác nhân giống và phục hồi rạn san hô. Trong đó, kỹ thuật vườn ươm dưới nước được xem là một phương pháp hiệu quả để bảo tồn và phục hồi các loài san hô tạo rạn quan trọng. Nghiên cứu này đánh giá tốc độ tăng trưởng và tỷ lệ sống của hai loài A. muricata và A. robusta sau gần một năm trồng phục hồi tại Trạm Nghiên cứu Biển Đầm Báy, Vịnh Nha Trang trên giá thể dây treo và cọc sắt trên rạn đá tự nhiên. Kết quả cho thấy, điều kiện môi trường tại khu vực nghiên cứu phù hợp cho việc trồng phục hồi, bảo tồn và duy trì vườn ươm san hô ở đây. Tỷ lệ sống của san hô trên giá thể dây treo đạt 68,09% (loài A. muricata), lên tới 100% (loài A. robusta); đối với giá thể cọc sắt trên rạn đá tự nhiên, tỷ lệ sống của 2 loài san hô thử nghiệm đều đạt 80%. Tốc độ tăng trưởng trên giá thể dây treo của loài A. muricata trung bình đạt 13,99 mm/tháng, loài A. robusta đạt 19,31 mm/tháng; đối với giá thể cọc sắt trên rạn đá tự nhiên, loài A. muricata có tốc độ tăng trưởng đạt 6,80 mm/tháng, loài A. robusta đạt 12,04 mm/tháng. Nghiên cứu này góp phần cung cấp dữ liệu thực nghiệm cho các chương trình phục hồi san hô, đồng thời mở ra hướng tiếp cận bền vững trong bảo tồn nguồn gen san hô tại Vịnh Nha Trang và các vùng biển lân cận.
Abstract
Growth and survival rate of two coral species of the acropora genus (A. muricata and A. robusta) on various settlement substrates at the Dam Bay marine research station, Nha Trang Bay
The severe decline in coral populations, particularly among Acropora species, has underscored the urgent need for coral propagation and reef restoration efforts. Among the available approaches, in situ nursery techniques are considered effective for conserving and rehabilitating essential reef-building coral species. This study evaluated the growth rate and survival of Acropora muricata and A. robusta after nearly one year of restoration at the Dam Bay Marine Research Station, Nha Trang Bay, using two substrate types: suspended ropes and iron stakes fixed to natural reef rock. The results indicated that the environmental conditions at the study site were suitable for coral nursery development and maintenance. On suspended rope substrates, survival rates reached 68.09% for A. muricata and up to 100% for A. robusta, whereas on iron stake substrates, both species exhibited a survival rate of 80%. The average growth rate of A. muricata and A. robusta on suspended ropes were 13.99 mm/month and 19.31 mm/month, respectively, whereas on iron stakes, the growth rates were 6.80 mm/month and 12.04 mm/month. These findings provide empirical evidence to support coral restoration programs and suggest a sustainable approach to conserving coral genetic resources in Nha Trang Bay and adjacent coastal areas.
Keywords: Coral reef; genus Acropora; growth, Nha Trang bay; restoration; survival rate.
Từ khóa
Bảo tồn, giống Acropora, rạn san hô, tăng trưởng, tỷ lệ sống, Vịnh Nha Trang
Chi tiết bài viết

Bài báo này được cấp phép theo Creative Commons Attribution 4.0 International License.
Điểm nổi bật:
Nghiên cứu cho thấy hai loài san hô Acropora muricata và A. robusta có tỷ lệ sống và tốc độ tăng trưởng cao khi được phục hồi bằng kỹ thuật vườn ươm (trên giá thể dây treo và cọc sắt) tại Đầm Báy, Vịnh Nha Trang.
Kết quả khẳng định tiềm năng ứng dụng của phương pháp này trong các chương trình phục hồi và bảo tồn san hô tạo rạn tại khu vực Vịnh Nha Trang và các vùng nước lân cận.
1. ĐẶT VẤN ĐỀ
Rạn san hô là một trong những hệ sinh thái biển có đa dạng sinh học cao nhất trên thế giới, đóng vai trò thiết yếu trong cân bằng sinh thái đại dương cũng như mang lại nhiều giá trị kinh tế, xã hội cho con người. Về mặt sinh thái, rạn san hô là nơi cư trú, sinh sản và cộng sinh của hàng nghìn loài sinh vật biển, hình thành nên các mạng lưới dinh dưỡng phức tạp và ổn định. Cấu trúc ba chiều của rạn còn đóng vai trò như “tường chắn sóng” tự nhiên, giúp giảm thiểu ảnh hưởng của bão lũ, hạn chế xói mòn bờ biển và góp phần vào quá trình tái tạo bãi biển. Bên cạnh đó, nhiều hợp chất sinh học có nguồn gốc từ san hô đã được nghiên cứu và ứng dụng trong y học, bao gồm các hợp chất có tiềm năng điều trị ung thư và các bệnh mãn tính khác [1, 2].
Tuy nhiên, trong vài thập kỷ qua, các rạn san hô trên toàn cầu đang đối mặt với sự suy thoái nghiêm trọng do tác động tổng hợp của biến đổi khí hậu, ô nhiễm môi trường, đánh bắt hủy diệt và các hoạt động ven biển thiếu bền vững. Tại Việt Nam, rạn san hô ở Vịnh Nha Trang - khu bảo tồn biển đầu tiên và tiêu biểu của quốc gia - cũng không nằm ngoài xu hướng này. Theo các báo cáo gần đây, độ phủ trung bình của san hô tại đây đã giảm tới 64,4%, trong đó hai giống san hô quan trọng là Acropora và Montipora lần lượt giảm tới 80,6% và 82,3% [3, 4]. Diện tích các rạn san hô khỏe mạnh tại khu vực này đã suy giảm mạnh từ 6,65 km² trước năm 1980 xuống chỉ còn 0,74 km² vào năm 2019 - tương đương mức suy giảm hơn 90%. Riêng tại khu vực Vịnh Nha Trang, diện tích rạn san hô đã giảm từ 275,72 ha vào năm 2002 xuống còn 84,34 ha vào năm 2024, tương đương với mức suy giảm 70% trong giai đoạn 22 năm [5].
Trước thực trạng đáng lo ngại này, nhiều quốc gia trong khu vực như Malaysia, Indonesia, Philippines, Singapore, Thái Lan và Việt Nam đã triển khai các chương trình phục hồi rạn san hô với các kỹ thuật đa dạng, bao gồm: di dời tập đoàn san hô, phân mảnh cấy ghép, và tạo giá thể nhân tạo phục vụ phục hồi [6]. Tại Việt Nam, các phương pháp phục hồi chủ yếu tập trung vào kỹ thuật tách mảnh san hô giống có kích thước tập đoàn >10 cm và cấy trực tiếp vào môi trường tự nhiên. Một số nghiên cứu điển hình như Võ Sĩ Tuấn và cộng sự (2009) tại Hòn Ngang - Hòn Nhàn (Bình Định), hay Hoàng Xuân Bền (2006) tại Vịnh Nha Trang đã cho thấy tỷ lệ sống của các mảnh san hô phục hồi đạt từ 80–100% tại các khu vực ít chịu tác động tiêu cực từ môi trường và con người [7, 8].
Trong số các loài san hô cứng, Acropora là giống có ý nghĩa đặc biệt quan trọng vì đóng vai trò nền tảng trong việc hình thành cấu trúc rạn, tạo môi trường sống và duy trì sự ổn định của hệ sinh thái san hô. Do đó, bảo tồn và phục hồi Acropora là ưu tiên hàng đầu trong các chương trình phục hồi rạn san hô hiện nay. Đặc biệt, trong bối cảnh biến đổi khí hậu và các áp lực môi trường ngày càng gia tăng, việc nghiên cứu khả năng thích nghi và tăng trưởng của Acropora sẽ góp phần làm sáng tỏ cơ chế chống chịu của rạn san hô nói chung trước các biến động môi trường [9, 10].
Mặc dù đã có một số nghiên cứu phục hồi san hô được thực hiện tại Việt Nam, nhưng vẫn còn thiếu các dữ liệu thực nghiệm chi tiết về tốc độ tăng trưởng và tỷ lệ sống của giống Acropora tại vùng biển Nha Trang. Nghiên cứu này nhằm đánh giá tốc độ sinh trưởng và tỷ lệ sống của các tập đoàn san hô giống Acropora sau một năm trồng phục hồi tại khu vực nghiên cứu. Kết quả sẽ cung cấp cơ sở khoa học quan trọng phục vụ công tác bảo tồn, lưu giữ nguồn gen và nâng cao hiệu quả phục hồi rạn san hô bền vững trong tương lai.
2. VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
Đối tượng: 2 loài san hô thuộc giống Acropora (A. muricata và A. robusta).
Địa điểm: Trạm Nghiên cứu và Thử nghiệm biển Đầm Báy, Vịnh Nha Trang (Hình 1).
Thời gian: Từ tháng 8/2024 đến tháng 7/2025.
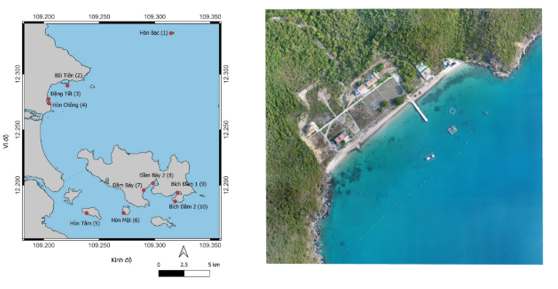
Hình 1. Bản đồ khu vực nghiên cứu tại vịnh Nha Trang, tỉnh Khánh Hòa. Các chấm màu đỏ biểu thị 10 điểm thu mẫu nghiên cứu, gồm: Hòn Bạc (1), Bãi Tiên (2), Đăng Tà (3), Hòn Chồng (4), Hòn Tằm (5), Hòn Một (6), Đầm Bấy (7), Đầm Bấy 2 (8), Đạn 1 (9) và Bích Đầm 2 (10). Ảnh vệ tinh bên phải thể hiện vị trí Trạm Nghiên cứu và Thử nghiệm biển Đầm Báy
Phương pháp thu thập giống san hô: 2 loài san hô được thu thập ở khu vực Đầm Báy, Vịnh Nha Trang (thu không quá 10% từ san hô nguồn). Các mảnh san hô này được thu từ ít nhất 20 tập đoàn nguồn cho mỗi loài nhằm đảm bảo tính đa dạng nguồn giống và giảm sai lệch do lấy từ cùng tập đoàn. San hô sau khi thu được chuyển ngay vào các xô nhựa chứa nước biển, đảm bảo luôn ở trạng thái ngập hoàn toàn trong quá trình vận chuyển. Các mảnh san hô sau đó được đặt vào rọ nhựa để thích nghi trong điều kiện tự nhiên tại điểm thí nghiệm trong 1–2 ngày trước khi tiến hành gắn lên giá thể. Các mảnh san hô từ mỗi tập đoàn nguồn sẽ được phân bổ đồng đều cho hai loại giá thể, đảm bảo mỗi giá thể nhận ≥ 1 mảnh từ mỗi nguồn san hô.
Phương pháp tạo giá thể và trồng thử nghiệm
Giá thể dây treo: Các dây cước đúc đường kính 2 mm, dài 5 m được cố định bằng hệ thống phao nổi và các dây neo theo chiều thẳng đứng cách nhau 80 cm. Vị trí tạo giá thể ở độ sâu 6 đến 7 m. Sau đó, các tập đoàn san hô được cố định vào các sợi dây cước bằng dây rút ở các vị trí có khoảng cách tính từ mặt nước (1 m; 1,5 m; 2 m; 2,5 m, 3 m; 3,5 m). Tổng cộng có 47 tập đoàn A. muricata với kích thước ban đầu Lo = 110,19 ± 11,90 mm và 46 tập đoàn A. robusta với kích thước Lo = 116,55 ± 34,31 mm được trồng và gắn nhãn để tiến hành theo dõi về tăng trưởng và tỷ lệ sống.
Giá thể cọc sắt: Các cọc sắt có đường kính 16 đến 18 mm được đóng cố định vững chắc trên rạn đá tự nhiên (độ sâu của rạn đá 2 - 4 m). Tương tự, các tập đoàn san hô được cố định bằng các dây rút trên các cọc sắt (mỗi cọc 1 tập đoàn), 50 tập đoàn A. muricata với kích thước Lo = 104,90 ± 16,59 mm và 50 tập đoàn A. robusta với kích thước Lo = 132,9 ± 23,79 mm được trồng và gắn nhãn để tiến hành theo dõi về tăng trưởng và tỷ lệ sống.
Phương pháp xác định một số thông số môi trường chính
Phương pháp xây dựng sơ đồ độ sâu tại Đầm Báy
Nội suy không gian là phương pháp được sử dụng để xây dựng bản đồ độ sâu [11]. Trong số các kỹ thuật nội suy không gian, phương pháp nội suy nghịch đảo khoảng cách có trọng số (IDW – Inverse Distance Weighted) được xem là một tiếp cận đơn giản, trực quan và dễ diễn giải, đồng thời vẫn đáp ứng độ chính xác và độ tin cậy cần thiết, và cũng đã được tích hợp phổ biến trong hầu hết các phần mềm GIS [12]. Do đó, phương pháp IDW được lựa chọn áp dụng để xây dựng bản đồ độ sâu tại Trạm Nghiên cứu và Thử nghiệm biển Đầm Báy.
Dữ liệu độ sâu tại khu vực Trạm Nghiên cứu và Thử nghiệm biển Đầm Báy được thu thập vào tháng 9/2024 bằng thiết bị đo sâu đơn tia GPSMAP 585 PLUS (GARMIN, Đài Loan), tổng cộng 186 điểm được thu thập. Giá trị độ sâu được nội suy bằng phương pháp nghịch đảo khoảng cách có trọng số (IDW), trong đó giá trị tại điểm cần nội suy được xác định dựa trên trung bình trọng số của các điểm lân cận [13]. Giá trị độ sâu cần nội suy tại vị trí x0, kí hiệu (x0) được xác định theo phương trình (1) [14], trong phương:
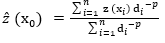
(1)
Trong đó:
z (xi) là giá trị của độ sâu tại vị trí được đo đạc i
n là số vị trí lấy mẫu lân cận được sử dụng cho ước tính (x0)
di là khoảng cách giữa điểm x0 và điểm xi
p là tham số lũy thừa, tham số p được thiết lập bằng 2.
Phương pháp xác định một số thông số môi trường: Các thông số môi trường biển (nhiệt độ, chỉ số pH, hàm lượng oxy hòa tan, độ mặn) được đo trực tiếp tại hiện trường bằng máy đo cầm tay pH HI 8314-1, máy đo oxy hòa tan Hanna HI 9147-4, máy đo độ mặn HI 98319. Trong quá trình đo, thả trực tiếp đầu đo của máy đo pH và oxy hòa tan (DO) vào trong nước biển, cho ngập hoàn toàn đầu đo (cách mặt nước từ 10 -15 cm), sau đó bật máy đo, chờ cho các giá trị pH và oxy hòa tan trên màn hình hiển thị ổn định, không dao động thì ghi lần lượt các kết quả đo được. Hiệu chuẩn thiết bị trước khi đo (máy đo pH hiệu chỉnh bằng dung dịch pH 7,01 HI 7007 và pH 10,01 HI 7010; máy đo hàm lượng oxy hòa tan hiệu chỉnh bằng dung dịch oxy zero HI 7040L; máy đo độ mặn hiệu chỉnh bằng dung dịch NaCl 35,00 g/l HI 70024P theo hướng dẫn của nhà sản xuất).
Phương pháp chăm sóc và thu thập số liệu
San hô được kiểm tra định kỳ hai lần mỗi tháng, kết hợp vệ sinh giá thể và ghi nhận tình trạng sức khỏe của các tập đoàn. Trong mỗi đợt kiểm tra, tiến hành loại bỏ toàn bộ ốc Drupella gây hại và dọn sạch các vật bám không mong muốn trên san hô và giá thể như túi nilon, dây cước, bao bì nhựa và các mảnh vụn khác, nhằm hạn chế cạnh tranh không gian, giảm stress cơ học và đảm bảo điều kiện phát triển ổn định cho san hô. Sử dụng máy Olympus TG6 để quay phim, chụp hình các tập đoàn san hô được trồng, cũng như các sinh vật gây hại và sinh vật cộng sinh khác. Trong quá trình chụp các tập đoàn san hô, máy ảnh được giữ ổn định, với ống kính đặt vuông góc với mặt phẳng mẫu nhằm hạn chế sai lệch do góc nhìn; thước chuẩn được bố trí cùng mặt phẳng với nhánh san hô. Khoảng cách giữa máy ảnh - mẫu - thước được duy trì không đổi nhằm đảm bảo tính nhất quán của dữ liệu hình ảnh. Các nhóm sinh vật cộng sinh và sinh vật gây hại sẽ được định danh bằng tham vấn các chuyên gia và một số tài liệu tham khảo thông dụng.
Tỷ lệ sống:
Tỷ lệ sống (%) = (N1/N0) × 100 (2)
Trong đó: N0 là số lượng tập đoàn san hô lưu giữ, N1 là số lượng san hô sống theo thời gian kiểm tra.
Tốc độ tăng trưởng được xác định theo công thức:
L (mm/tháng) = (Lt – L0)/(tt) (3)
Trong đó: Lt là chiều dài ở các thời gian đo (mm); L0 là chiều dài ở thời gian bắt đầu thí nghiệm; tt là các thời gian đo (tháng). Phần mềm ImageJ được sử dụng để đo chiều dài từ các ảnh chụp, với quy trình hiệu chuẩn kích thước thực hiện trực tiếp trên từng ảnh dựa vào độ dài thực của thước chuẩn. Chiều dài Lt của san hô được đo ở kích thước lớn nhất từ điểm đầu tới điểm cuối của tập đoàn san hô, sai số đo bằng phần mềm ImageJ ước lượng khoảng ± 0,6 mm.
Phương pháp xử lý số liệu: dữ liệu được phân tích và xử lý bằng ngôn ngữ lập trình Python (phiên bản 3.10).
3. KẾT QUẢ
3.1. Các thông số môi trường chính tại Trạm Đầm Báy
Bản đồ độ sâu tại khu vực xung quanh Trạm Nghiên cứu và Thử nghiệm biển Đầm Báy cho thấy khu vực này có độ sâu tương đối thấp, dao động từ 0 đến -17,8 m, với giá trị độ sâu trung bình -6,5 m. Đặc biệt, phần mặt nước nằm trong ranh giới của Trạm Nghiên cứu và Thử nghiệm biển Đầm Báy có độ sâu trung bình -4,1 m, với giá trị độ sâu thay đổi từ 0 đến -7,6 m. Điều này tạo điều kiện thuận lợi cho sự sinh tồn và phát triển của san hô trong khu vực ranh giới của Trạm (Hình 2).
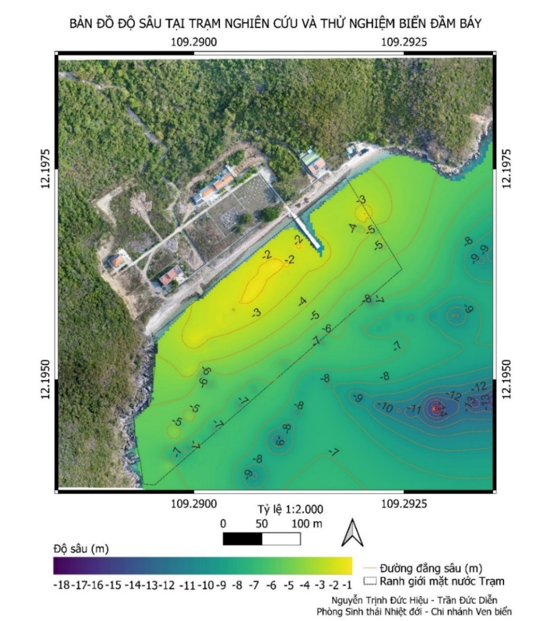
Hình 2. Bản đồ độ sâu tại Trạm Nghiên cứu và Thử nghiệm biển Đầm Báy
Kết quả đánh giá độ chính xác mô hình nội suy IDW cho thấy sự tương quan cao giữa giá trị độ sâu dự báo và giá trị đo đạc thực tế (Hình 3). Mô hình đạt RMSE = 1,38 m và hệ số xác định R² = 0,93, phản ánh khả năng giải thích biến thiên dữ liệu tốt và sai số dự báo thấp. Điều này khẳng định phương pháp IDW cho hiệu quả ước lượng độ sâu đáng tin cậy tại khu vực nghiên cứu.
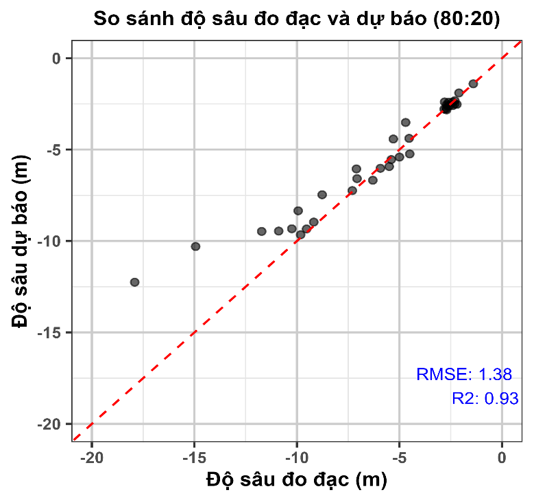
Hình 3. Kiểm định độ sâu đo đạc và độ sâu dự báo từ mô hình
Các thông số môi trường tại Trạm Nghiên cứu và Thử nghiệm biển Đầm Báy từ tháng 6/2024 đến tháng 6/2025 được thể hiện ở Hình 4. Các thông số môi trường như nhiệt độ, độ mặn, pH, oxy hòa tan phù hợp cho quá trình trồng phục hồi và lưu giữ nguồn gen thuộc giống Acropora. Nhiệt độ trung bình khoảng 27,8oC, tuy nhiên vào tháng 9/2024 ghi nhận được nhiệt độ bề mặt lớn hơn 30oC. Độ mặn cao trung bình đạt 32,6‰; giá trị pH là 8,15; oxy hòa tan là 7,4 mg/l.
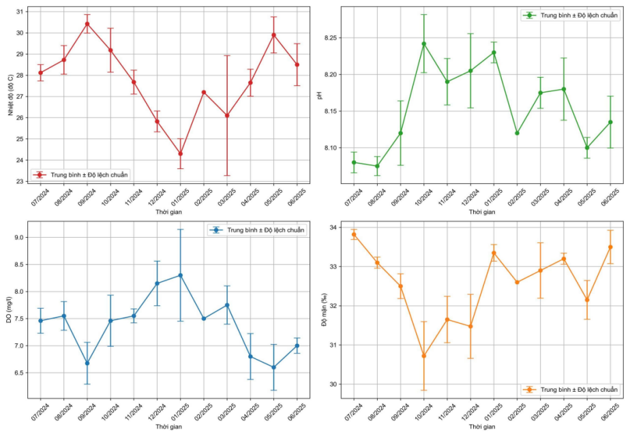
Hình 4. Một số thông số môi trường tại Trạm Nghiên cứu và Thử nghiệm biển Đầm Báy trong thời gian tiến hành thí nghiệm
3.2. Tỷ lệ sống của san hô
Sau gần một năm trồng thử nghiệm tỷ lệ sống của san hô ở khu vực vườn ươm thuộc Trạm nghiên cứu và Thử nghiệm biển Đầm Báy khá cao được thể hiện ở Hình 5. Đối với giá thể dây treo theo chiều thẳng đứng, loài san hô A. muricata đạt 68,09%, trong khi loài A. robusta tỉ lệ sống đạt 100%. Đối với giá thể cọc sắt trên rạn đá tự nhiên, tỷ lệ sống của 2 loài san hô thử nghiệm đều đạt 80 % (Hình 5, Hình 6).
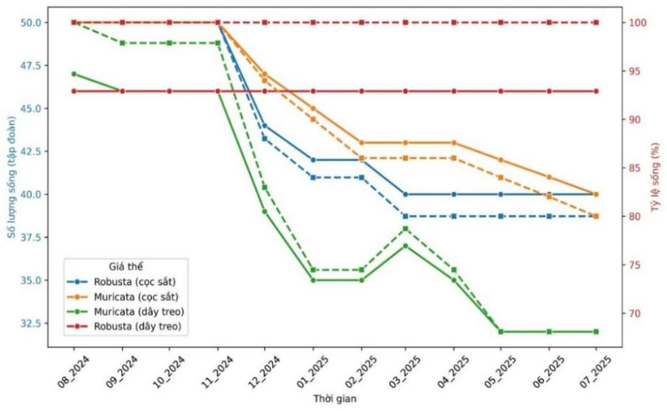
Hình 5. Biến động số lượng tập đoàn (trục tung bên trái) và tỷ lệ sống (trục tung bên phải) của san hô giống thuộc hai loài A. robusta và A. muricata trên hai dạng giá thể gồm cọc sắt và dây treo trong thời gian theo dõi từ tháng 8/2024 đến tháng 7/2025. Đường liền biểu thị số lượng tập đoàn san hô còn sống; đường nét đứt biểu thị tỷ lệ sống (%). Màu xanh dương: A. robusta trên cọc sắt; màu cam: A. muricata trên cọc sắt; màu xanh lá cây: A. muricata trên dây treo; màu đỏ: A. robusta trên dây treo.

Hình 6. Các giá thể phục hồi san hô (dây treo và cọc sắt trên rạn đá tự nhiên) lúc bắt đầu và kết thúc thí nghiệm
3.3. Tăng trưởng của san hô
Sau gần một năm trồng thực nghiệm, cho thấy sự gia tăng đáng kể về tốc độ tăng trưởng của hai loài san hô (Hình 7). Trên giá thể dây treo thẳng đứng, A. muricata đạt tốc độ tăng trưởng theo tháng trung bình là 13,99 mm/tháng, trong khi, A. robusta ghi nhận mức tăng trưởng cao hơn, đạt 19,31 mm/tháng. Trên giá thể cọc sắt cố định tại rạn đá tự nhiên, tốc độ tăng trưởng trung bình của A. muricata đạt 6,80 mm/tháng và của A. robusta cao hơn đạt 12,04 mm/tháng ((Hình 7, Hình 8, Hình 9, Hình 10, Hình 11).
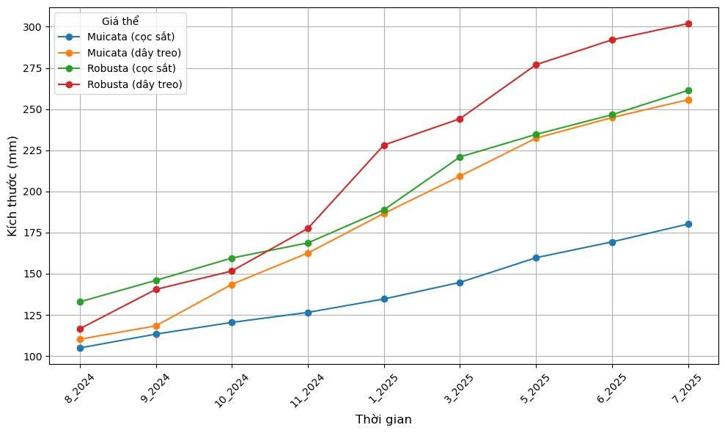
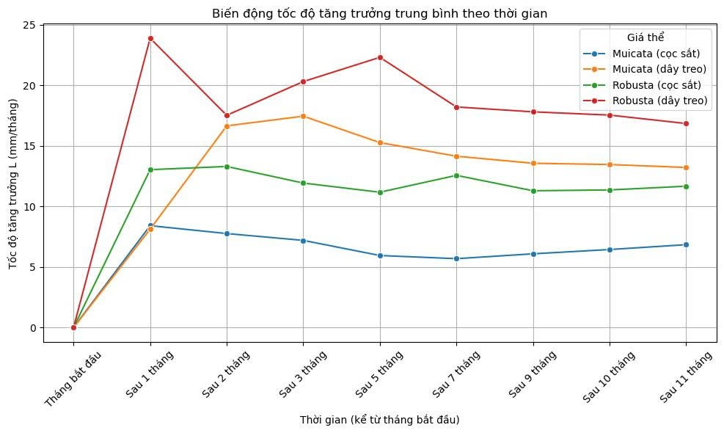
Hình 7. Tăng trưởng và biến động tốc độ tăng trưởng theo thời gian của các loài san hô ở các giá thể khác nhau
 Bắt đầu (tháng 8/2024) |  Sau 1 tháng (9/2024) |  Sau 2 tháng (10/2024) |
 Sau 3 tháng (11/2024) |  Sau 7 tháng (3/2025) |  Sau 9 tháng (5/2025) |
 Sau 10 tháng (6/2025) |  Sau 11 tháng (7/2025) |
Hình 8. Tăng trưởng của loài A. muricata (mã số M18) ở giá thể cọc sắt trên rạn đá tự nhiên sau các tháng trồng phục hồi
 Bắt đầu (tháng 8/2024) |  Sau 1 tháng (9/2024) |  Sau 2 tháng (10/2024) |
 Sau 3 tháng (11/2024) |  Sau 7 tháng (3/2025) |  Sau 9 tháng (5/2025) |
 Sau 10 tháng (6/2025) |  Sau 11 tháng (7/2025) |
Hình 9. Tăng trưởng của loài A. robusta (mã số R29) ở giá thể cọc sắt trên rạn đá tự nhiên sau các tháng trồng phục hồi
 Bắt đầu (tháng 8/2024) |  Sau 1 tháng (tháng 9/2024) |
 Sau 2 tháng (tháng 10/2024) |  Sau 3 tháng (tháng 11/2024) |
 Sau 7 tháng (tháng 3/2025) |  Sau 9 tháng (tháng 5/2025) |
 Sau 10 tháng (tháng 6/2025) |  Sau 11 tháng (tháng 7/2025) |
Hình 10. Tăng trưởng của loài A. muricata (mã số M18) trên giá thể dây treo sau các tháng trồng phục hồi
 Bắt đầu (tháng 8/2024) |  Sau 1 tháng (tháng 9/2024) |
 Sau 2 tháng (tháng 10/2024) |  Sau 3 tháng (tháng 11/2024) |
 Sau 7 tháng (tháng 3/2025) |  Sau 9 tháng (tháng 5/2025) |
 Sau 10 tháng (tháng 6/2025) |  Sau 11 tháng (tháng 72025) |
Hình 11. Tăng trưởng của loài A. muricata (mã số R35) trên giá thể dây treo sau các tháng trồng phục hồi
4. THẢO LUẬN
Sự thành công của phương pháp trồng phục hồi san hô giống Acropora phụ thuộc mạnh mẽ vào các yếu tố môi trường như cường độ ánh sáng, nhiệt độ nước, độ mặn, động lực dòng chảy, và chất lượng nước (bao gồm hàm lượng chất dinh dưỡng, độ trong và mức độ lắng đọng trầm tích). Các nghiên cứu chỉ ra rằng nhiệt độ lý tưởng cho Acropora phát triển dao động từ 26–29°C [15]; ngưỡng nhiệt độ gây tẩy trắng san hô xác định cho khu vực Nam Việt Nam là 30°C [16]. Ánh sáng mặt trời đầy đủ là yếu tố cần thiết cho quá trình cộng sinh với zooxanthellae, nhưng bức xạ UV cao hoặc nước đục sẽ làm giảm hiệu suất quang hợp. Dòng chảy ổn định vừa phải giúp cung cấp oxy và dinh dưỡng, đồng thời loại bỏ trầm tích khỏi bề mặt san hô non. Ngoài ra, ô nhiễm từ đất liền và phát triển ven bờ cũng là những tác nhân gây ức chế hoặc gây chết cho các tập đoàn san hô phục hồi. Chất lượng môi trường tại Trạm Nghiên cứu và Thử nghiệm biển Đầm Báy được xem là phù hợp, tạo điều kiện thuận lợi cho sự sinh trưởng và phát triển của san hô. Tuy nhiên, hiện tượng nóng lên toàn cầu dẫn tới nhiệt độ nước bề mặt ở Đầm Báy có tháng vượt ngưỡng 30°C, có thể làm ảnh hưởng tới quá trình sinh trưởng và gây nên hiện tượng tẩy trắng của san hô được phục hồi.
Trong nghiên cứu này, chúng tôi không sử dụng các kiểm định thống kê, vì kích thước san hô trồng đầu vào khác nhau, điều này có thể ảnh hưởng tới tỷ lệ sống và tăng trưởng của chúng khi tiến hành nuôi thử nghiệm. Theo Edwards & Gomez (2007), tỷ lệ sống của Acropora bị ảnh hưởng theo kích thước, cách ghép và điều kiện môi trường sau khi cấy ghép. Các mảnh (tập đoàn) san hô lớn hơn (>5–10 cm) có tỷ lệ sống cao hơn đáng kể so với các mảnh nhỏ (2-3 cm) [17]. Theo Lirman và cộng sự (2014), tốc độ tăng trưởng của A. cervicornis trung bình: 20–173 cm/năm, tùy theo kích thước ban đầu và điều kiện, tăng trưởng cao nhất được ghi nhận ở các mảnh nhỏ (<15 cm): ~59–89 cm/năm tại vườn ươm [18]. Theo Khasanah và cộng sự (2020), tốc độ tăng trưởng và tỷ lệ sống của một số loài san hô giống Acropora tại Paiton, Indonesia: A. formosa – A.muricata có tốc độ tăng trưởng đạt 1,958 cm/tháng và tỷ lệ sống sau 5 tháng là 82,61%; A. intermedia là 1,730 cm/tháng, tỷ lệ sống sau 5 tháng là 78,26%; A. pulchra là 0,958 cm/tháng, tỷ lệ sống sau 5 tháng là 80,95%; A. gomezi là 0,756 cm/tháng, tỷ lệ sống sau 5 tháng là 73,91% [19]. Tại vùng biển Đầm Báy, theo Vũ Việt Dũng và cộng sự (2024), tỷ lệ sống của san hô phục hồi trên reefball đạt từ 64,7 đến 79,4%, trung bình đạt 69,4%; tỷ lệ sống lớn nhất thuộc về loài Pocillopora verruscosa 79,4% và thấp nhất là loài A. formosa -A. muricata là 64,7%. Tốc độ sinh trưởng của loài A. nobilis trung bình 7,7 mm/tháng, loài A. formosa – A. muricata trung bình 6,7 mm/tháng [20]. Theo nghiên cứu này, tỷ lệ sống của san hô giống Acropora trồng ở vườn ươm Đầm Báy cao từ 68,06% đến 100%, trung bình là 80% trên giá thể dây treo và giá thể cọc sắt. Tốc độ tăng trưởng trên giá thể dây treo, loài A. muricata có trung bình đạt 13,99 mm/tháng, trong khi loài A. robusta có tốc độ tăng trưởng đạt 19,31 mm/tháng. Đối với giá thể cọc sắt trên rạn đá tự nhiên, loài A. muricata có tốc độ tăng trưởng đạt 6,80 mm/tháng, trong khi loài A. robusta có tốc độ tăng trưởng đạt 12,04 mm/tháng. Kết quả này cao hơn so với số liệu trước đây tại Đầm Báy do Vũ Việt Dũng và cộng sự (2024) ghi nhận, trong đó tốc độ tăng trưởng của A. formosa – A. muricata trung bình là 6,7 mm/tháng và A. nobilis là 7,7 mm/tháng trên reefball. Sự khác biệt này có thể được lý giải bởi một số yếu tố. Thứ nhất, nghiên cứu của chúng tôi sử dụng giá thể dây treo theo phương thẳng đứng và cọc sắt cố định, giúp hạn chế lắng đọng trầm tích và tạo điều kiện lưu thông nước tốt hơn so với reefball, từ đó tăng cường cung cấp oxy và dinh dưỡng cho san hô. Bên cạnh đó, điều kiện môi trường tại thời điểm thí nghiệm (2024–2025) tại Trạm nghiên cứu và Thử nghiệm biển Đầm Báy nhìn chung ổn định và thuận lợi cho sinh trưởng của san hô, góp phần nâng cao tốc độ tăng trưởng. Do vậy, mặc dù tăng trưởng và tỷ lệ sống cao hơn so với các nghiên cứu trước, nhưng vẫn phản ánh hợp lý ảnh hưởng của loài, loại giá thể, và điều kiện môi trường đến hiệu quả phục hồi san hô. Phương pháp trồng phục hồi này cao hơn so với trồng trên reefball và phù hợp với tăng trưởng của các loài san hô giống Acropora được báo cáo tại Indonesia. Nhìn chung, vùng biển thuộc Trạm Nghiên cứu và Thử nghiệm biển Đầm Báy, Vịnh Nha Trang phù hợp để bảo tồn, lưu giữ và trồng phục hồi các loài san hô cứng quan trọng thuộc giống Acropora.
Địch hại và sinh vật cộng sinh với san hô
Sau một tháng trồng phục hồi, đã xuất hiện nhóm ốc Drupella gây hại cho san hô. Số lượng ốc này được loại trừ sau các đợt chăm sóc và vệ sinh san hô. Các nghiên cứu trước đây đã chỉ ra, ốc Drupella là một trong những loài động vật thân mềm ăn san hô có ảnh hưởng tiêu cực rõ rệt đến các rạn san hô ở khu vực Ấn Độ Dương - Thái Bình Dương. Các loài Drupella, đặc biệt là Drupella cornus (Hình 12) chủ yếu ăn các loài san hô cứng thuộc họ Acroporidae và Pocilloporidae. Chúng dùng bộ răng mài để cạo mô san hô sống, để lại bộ xương trắng trơ trọi, khiến san hô dễ bị nhiễm bệnh và giảm khả năng tái sinh. Khi mật độ Drupella tăng cao, chúng có thể tạo ra các vết thương trên diện rộng, góp phần làm suy giảm cấu trúc rạn và đa dạng sinh học của hệ sinh thái. Do đó, trong quá trình phục hồi rạn cần thường xuyên loại bỏ nhóm ốc này.
 |  |
Hình 12. Mặt trên và mặt dưới của ốc địch hại Drupella cornus
Khi rạn san hô được phục hồi, đã ghi nhận sự gia tăng đáng kể về số lượng và đa dạng loài cá. Sự tái sinh của các rạn san hô, đặc biệt là các loài san hô tạo rạn như Acropora, đã cung cấp lại nơi cư trú, sinh sản và nguồn thức ăn cho nhiều loài cá rạn. Các nhóm cá rạn đã xuất hiện với mật độ cao hơn, trong đó nhiều loài có giá trị sinh thái và kinh tế, cho thấy sự hồi phục của chuỗi thức ăn và sự ổn định của môi trường sống (Hình 13). Kết quả khảo sát ban đầu cho thấy sự gia tăng đáng kể về cả thành phần loài và mật độ cá rạn san hô tại khu vực biển thuộc trạm Đầm Báy, với hơn 256 loài cá được ghi nhận trong quá trình theo dõi.
 Canthigaster valentini |  Thalassoma lunare |
Hình 13. Một số loài cá xuất hiện ở rạn san hô trồng thử nghiệm
Các nhóm thân mềm bao gồm thân mềm hai mảnh vỏ (Bivalvia), ốc ký cư (Vermetidae), ốc xoắn (Gastropoda), thường bám vào san hô, đóng vai trò trong lưới thức ăn duy trì các quần xã sinh vật cộng sinh (Hình 14). Bên cạnh đó, cũng ghi nhận sự xuất hiện của các nhóm sinh vật khác như Trai trai tượng, cầu gai, nhum sọ tại rạn san hô trên nền đá được phục hồi. Nhìn chung, các nhóm thân mềm cộng sinh hoặc sống trong rạn san hô đóng vai trò sinh thái rõ rệt: từ lọc nước, quang hợp cộng sinh, đến kiểm soát tảo và tạo cấu trúc vi mô cho các sinh vật nhỏ trú ẩn. Việc bảo tồn và hiểu rõ chức năng của chúng là điều cần thiết trong quản lý và phục hồi rạn san hô. Những nghiên cứu về sự hình thành quần xã sinh vật cộng sinh sau khi rạn san hô được phục hồi cần được nghiên cứu thêm để góp phần bảo vệ rạn và duy trì sự phát triển ổn định của các quần xã sinh vật sau khi được hình thành.
 |  |
 |  |
 |  |
 |  |
Hình 14. Một số nhóm thân mềm cộng sinh cùng rạn san hô được phục hồi
Kết quả ban đầu cho thấy, quần xã sinh vật ổn định hình thành quanh các cụm san hô phục hồi sẽ góp phần duy trì cân bằng sinh thái, tăng khả năng tự phục hồi của hệ sinh thái biển và nâng cao sức chống chịu trước biến đổi khí hậu và tác động của con người. Đây là nền tảng quan trọng để bảo vệ đa dạng sinh học và đảm bảo nguồn lợi thủy sản bền vững cho cộng đồng ven biển.
5. KẾT LUẬN
Trạm Nghiên cứu và Thử nghiệm biển Đầm Báy có độ sâu trung bình 4,1 m, với giá trị độ sâu thay đổi từ 0 đến 7,6 m. Các thông số môi trường cơ bản như pH, DO, độ mặn phù hợp cho việc phục hồi và lưu giữ nguồn gen các loài san hô thuộc giống Acropora (A. muricata, A. robusta).
Tỷ lệ sống của loài A. muricata đạt 68,09%, trong khi loài A. robusta tỉ lệ sống đạt 100% trên giá thể dây treo. Đối với giá thể cọc sắt trên rạn đá tự nhiên, tỷ lệ sống của 2 loài san hô thử nghiệm đều đạt 80%.
Tốc độ tăng trưởng trên giá thể dây treo, loài A. muricata có trung bình đạt 13,99 mm/tháng, trong khi loài A. robusta có tốc độ tăng trưởng đạt 19,31 mm/tháng. Đối với giá thể cọc sắt trên rạn đá tự nhiên, loài A. muricata có tốc độ tăng trưởng đạt 6,80 mm/tháng, trong khi loài A. robusta có tốc độ tăng trưởng đạt 12,04 mm/tháng. Nhìn chung, trên giá thể dây treo, san hô có tốc độ tăng trưởng cao hơn trên giá thể cọc sắt, và loài A. robusta có tốc độ tăng trưởng cao hơn loài A. muricata.
Tóm lại, ưu tiên sử dụng giá thể dây treo cho giai đoạn nhân giống, phân mảnh san hô Acropora, trong khi cọc sắt trên rạn tự nhiên thích hợp cho giai đoạn phục hồi rạn lâu dài; thường xuyên vệ sinh và loại bỏ địch hại như ốc Drupella, rác thải định kỳ 2 tuần/lần; thiết lập ngưỡng cảnh báo nhiệt ≥ 30–31°C kéo dài > 7 ngày để giám sát tẩy trắng; đồng thời mở rộng mô hình và đa dạng hóa loài san hô cần phục hồi.
Lời cảm ơn: Cảm ơn Bộ Quốc phòng đã cấp kinh phí thực hiện theo Chương trình bảo tồn nguồn gen năm 2024 cho nhiệm vụ “Lưu giữ, bảo tồn nguồn gen một số loài san hô thuộc giống Acropora tại vùng biển Đầm Báy, vịnh Nha Trang” (Quyết định số 3268/QĐ-TTNĐVN ngày 13/8/2024). Chúng tôi gửi lời cảm ơn chân thành tới các đồng nghiệp từ Viện Hàn lâm Khoa học Nga là Tiến sĩ Gorin S.A., Mekhova E.S đã hỗ trợ định danh các nhóm sinh vật cộng sinh và địch hại.
Tuyên bố về việc sử dụng Gen AI: Nhóm tác giả xin khẳng định rằng không sử dụng bất kỳ công cụ GenAI nào để tạo dữ liệu, xử lý số liệu, phân tích kết quả hoặc xây dựng các kết luận khoa học trong nghiên cứu này. Toàn bộ dữ liệu, kết quả phân tích và nội dung khoa học của bài báo được thực hiện trực tiếp bởi các tác giả và chịu trách nhiệm hoàn toàn về tính chính xác, trung thực học thuật của công trình.
Tuyên bố về đóng góp của tác giả: Trần Đức Diễn, Samoilov K.Yu., Nguyễn Trịnh Đức Hiệu, Đặng Đỗ Hùng Việt: Lên ý tưởng và thiết kế nghiên cứu; Trần Đức Diễn, Samoilov K.Yu., Dudka K.K., Nguyễn Trịnh Đức Hiệu, Đặng Đỗ Hùng Việt, Đồng Văn Kiên: Khảo sát thực địa và thu thập dữ liệu; Trần Đức Diễn, Nguyễn Trịnh Đức Hiệu, Hoàng Thị Ngọc Anh, Nguyễn Phương Liên: Phân tích dữ liệu, viết bản thảo bài báo; Trần Đức Diễn, Nguyễn Trịnh Đức Hiệu: Giám sát nghiên cứu, tìm kiếm nguồn kinh phí và hiệu chỉnh bản thảo cuối cùng để xuất bản.
Tuyên bố về xung đột lợi ích: Các tác giả không có xung đột về lợi ích.
Tài liệu tham khảo
- Status of Coral Reefs of the World 2020: 2021. http://www.unep.org/resources/status-coral-reefs-world-2020. [Accessed: 27/02/2024]
- Corals of Australia and the Indo-Pacific: 2020. https://uhpress.hawaii.edu/title/corals-of-australia-and-the-indo-pacific/. [Accessed: 13/05/2024]
- K. S. Tkachenko, N. H. Huan, N. H. Thanh and T. A. Britayev, Extensive coral reef decline in Nha Trang Bay, Vietnam: Acanthaster planci outbreak: the final event in a sequence of chronic disturbances, Mar. Freshw, Res., Vol. 72, pp.186–199, 2020. DOI:10.1071/MF20005
- K. S. Tkachenko, Coral reefs degradation under complex impact of natural and anthropogenic factors with Nha Trang Bay (Vietnam) as an example, Biol. Bull. Rev, Vol. 13, No. 5, pp. 442–459, 2023. DOI: 10.1134/S2079086423050079
- N. T. D. Hieu et al., Trends in coral reef habitats over two decades: lessons learned from Nha Trang Bay marine protected area, Vietnam, Water 2025, Vol.17, pp.1224, 2025. DOI: 10.3390/w17081224
- L. M. Chou, T. Yeemin, A. R. B. G. Yaman, S. T. Vo and P. Alino, Coral reef restoration in the South China Sea. Galaxea, Journal of Coral Reef Studies, Vol. 11, pp. 67-74, 2009.
- Võ Sĩ Tuấn, Nguyễn Xuân Hòa, Phan Kim Hoàng và Hoàng Xuân Bền, Phục hồi và quản lý rạn san hô ở nam vịnh Quy Nhơn, Bình Định, Tạp chí Khoa học và Công nghệ biển, Vol. IX, No.2, 2009.
- Hoàng Xuân Bền et al., Đánh giá sự ảnh hưởng của việc cắt cành các tập đoàn san hô trong quá trình phục hồi rạn san hô. Báo cáo tổng kết đề tài cơ sở, 2006.
- Genus Acropora: Implications of new biological findings for species concepts, Annual Review of Ecology and Systematics, Vol. 25, pp. 237–262, 2003.
- N. L. Rosser, L. Thomas, S. Stankowski, Z. T. Richards, W. J. Kennington and M. S. Johnson, Phylogenomics provides new insight into evolutionary relationships and genealogical discordance in the reef-building coral genus Acropora, Proceedings of the Royal Society B: Biological Sciences, Vol. 284, No. 1846, pp. 20162182, 2017.
- M. Curtarelli, J. Leão, I. Ogashawara, J. Lorenzzetti, and J. Stech. Assessment of spatial interpolation methods to map the bathymetry of an Amazonian hydroelectric reservoir to aid in decision making for water management, ISPRS International Journal of Geo-Information, Vol. 4, No. 1, p. 220-235, 2015.
- W. Maleika, Inverse distance weighting method optimization in the process of digital terrain model creation based on data collected from a multibeam echosounder, Applied Geomatics, Vol. 12, No. 4, pp. 397-407, 2020.
- S. Respati, T. Sulistyo, The effect of the number of inputs on the spatial interpolation of elevation data using IDW and ANNs, Geodesy and Cartography, Vol. 49, No. 1, pp. 60–65, 2023.
- J. Li, A. D. Heap, A review of spatial interpolation methods for environmental scientists, Geoscience Australia, 2008.
- C. Manullang et al., Responses of branching reef corals Acropora digitifera and Montipora digitata to elevated temperature and pCO2, PeerJ. 2020 Dec 21; Vol. 8, p. e10562, 2020. DOI: 10.7717/peerj.10562
- K. S. Tkachenko, V. V. Dung, V. T. Ha and H. H. Phong, Unprecedented coral bleaching in Southern Vietnam in 2024: the only hope for thermal refuges, Coral Reef, 2025. DOI: 10.1007/s00338-025-02651-8
- A. J. Edwards, E. D. Gomez, Reef Restoration Concepts and Guidelines: Making sensible management choices in the face of uncertainty. Coral Reef Targeted Research & Capacity Building for Management Programme, University of Queensland, 2007.
- D. Lirman et al., Growth dynamics of the threatened caribbean staghorn coral Acropora cervicornis: Influence of host genotype, symbiont identity, colony size, and environmental setting, PLoS ONE, Vol. 9, No. 9, pp. e107253, 2014. DOI: 10.1371/journal.pone.0107253
- R. I. Khasanah et al., Growth rate and survivorship of Acropora sp. fragments that transplanted on the artificial substrate made from fly ash and bottom ash, IOP Conference Series: Earth and Environmental Science, Vol. 441, pp. 012126, 2020. DOI: 10.1088/1755-1315/441/1/012126
- Vũ Việt Dũng, Trần Văn Bằng và Võ Thị Hà, Thử nghiệm phục hồi san hô trên các giá thể reefball tại khu vực Đầm Báy, vịnh Nha Trang, Tạp chí Khoa học và Công nghệ Nhiệt đới, số. 34 -06, 2024. DOI:10.58334/vrtc.jtst.n34.06